理化学研究所(理研)と科学技術振興機構(JST)は6月10日、毛包の幹細胞が従来の定説とは異なる細胞に由来し、既知のメカニズムとは別の仕組みで誘導されることを明らかにしたこと、ならびに毛包を構成する細胞の区画化と幹細胞誘導を同時に可能とする新しい形態形成モデル「テレスコープモデル」を提唱したことを発表した。
-
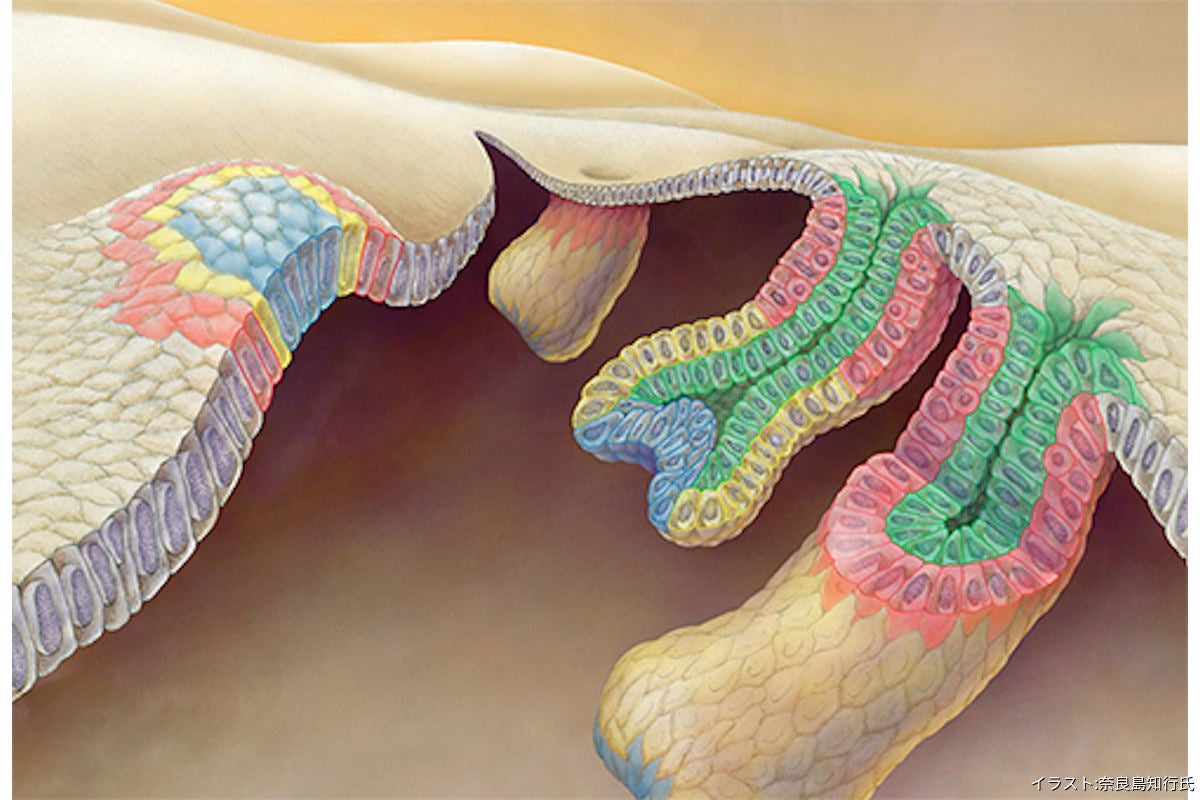
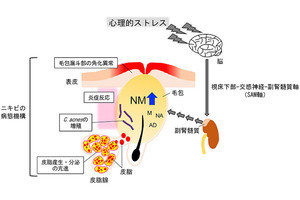
毛包幹細胞の発生起源(赤)と、今回提唱された新たな毛包発生モデル「テレスコープモデル」のイメージ イラスト:奈良島知行氏 (出所:理研Webサイト)

同成果は、理研 生命機能科学研究センター 細胞外環境研究チームの森田梨津子研究員、同・藤原裕展チームリーダーら13名が参加した研究チームによるもの。詳細は、英科学誌「Nature」にオンライン掲載された。
毛を産生する皮膚付属器官である毛包は、頭部をはじめ全身に分布し、身体の保護、感覚受容、容姿決定などの機能を担っているが、生涯にわたって「周期的に再生を繰り返す」ことから、いち早く成体における組織幹細胞(毛包幹細胞)の存在が特定された器官としても知られる。
しかし、マウスの成体毛包における毛包幹細胞の働きが明らかになる一方で、胚の発生過程において、一見すると均一な胎仔性の上皮細胞の集団から、どのようにして毛包幹細胞が正しい場所に正しいタイミングで誘導されるのかについてはよくわかっていなかったという。
2016年に米国の研究チームが、マウスの成体毛包幹細胞マーカー遺伝子の発現を指標とした遺伝学的細胞系譜解析に基づき、「毛包幹細胞の起源は、プラコード基底細胞の非対称分裂から生まれる転写因子SOX9陽性の基底上層細胞」と報告し、現在まで、この報告が定説として広く信じられてきたという。
今回の研究では、研究開始当時、未成熟な胎仔性上皮細胞の中で「毛包幹細胞になる細胞」を特異的に標識し得るマーカー遺伝子は同定されておらず、これが毛包幹細胞の起源や誘導メカニズムの理解が進まない最大の要因だと考えられたことから、既存の成体幹細胞マーカーに依存しない新しい手法として、毛包の発生を1細胞解像度で経時観察できる長期ex vivoライブイメージングを考案。さらに、1細胞トランスクリプトーム解析を組み合わせたマルチオミクスデータの統合的解析を実施することで、発生期毛包における「個々の毛包細胞の三次元的な位置や動態」、「細胞の発生系譜」、「各細胞系譜の遺伝子発現の変化」の解析に成功したという。
-
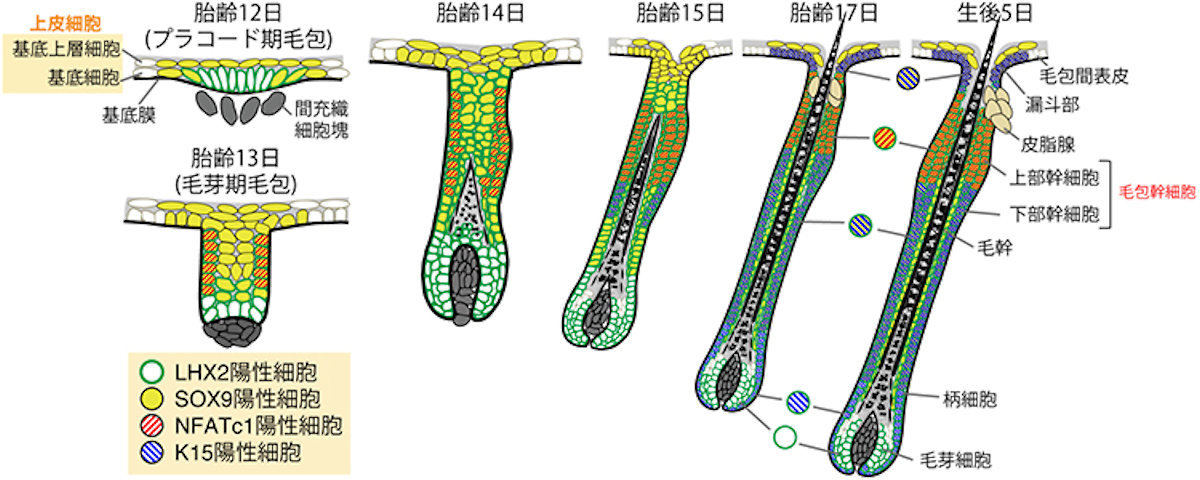
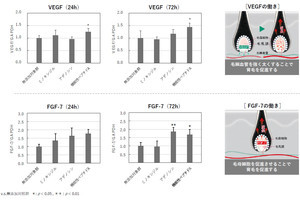
マウスの毛包発生の模式図。毛包は、上皮が内部に陥入してできた筒状の構造を取る。胎仔期に形成された毛包は、毛包幹細胞の働きにより生涯にわたって退縮と再生を繰り返すことが可能だ。成体の毛包幹細胞は、SOX9やLHX2といったマーカー遺伝子の発現によりほかの細胞と区別できるが、これらの遺伝子は発生過程のさまざまな細胞で発現するため、成体幹細胞マーカー遺伝子を頼りに幹細胞の発生起源を追跡することは困難であると考えられたという (出所:理研Webサイト)

-
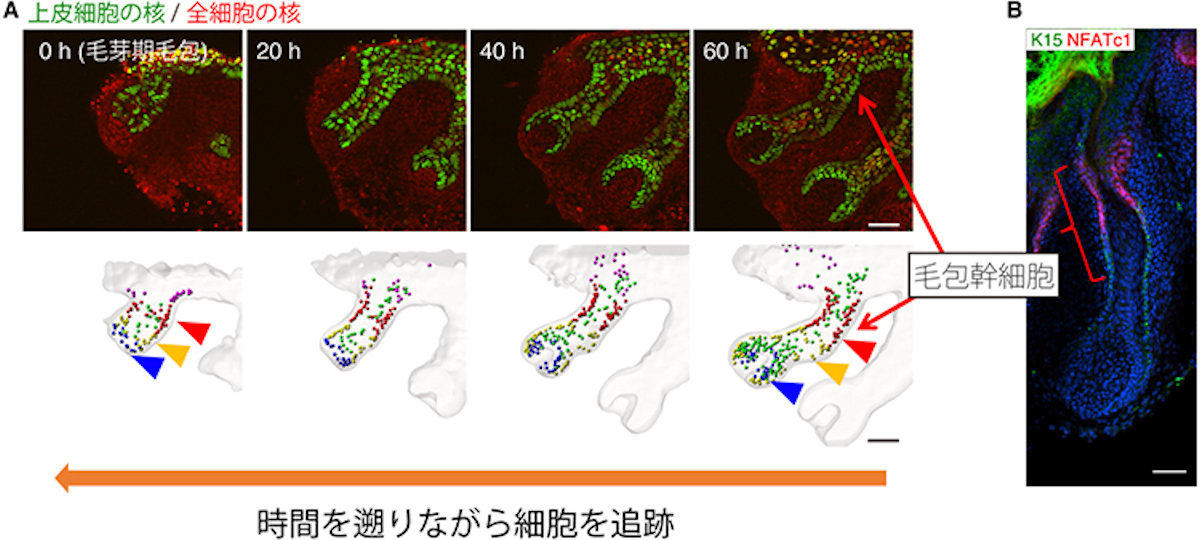
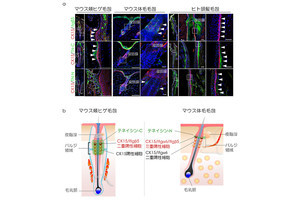
毛包発生の長期ex vivoライブイメージングと毛包幹細胞の発生系譜の同定。(A)マウスの胎齢13日毛芽期毛包から毛球性毛杭期(毛包が杭状の細長い形態を獲得する時期)のライブイメージング(上)と上皮細胞の遡及的追跡結果(下)。将来幹細胞になる細胞(赤矢頭)は毛芽期毛包の上部に存在し、他の細胞区画(黄、青)に移動しない。スケールバーは100μm。(B)生体外で発生した毛包の成体毛包幹細胞マーカー(緑、赤)での免疫染色像。スケールバーは50μm (出所:理研Webサイト)

具体的には、毛包発生の開始期の観察の結果、毛包プラコード(二次元の上皮シート構造)には、異なる系譜の細胞群が同心円リング状に配置されており、将来幹細胞になる細胞は、毛包プラコード辺縁に位置するSOX9陽性基底細胞のリングから生まれることが明らかとなったとする。
-
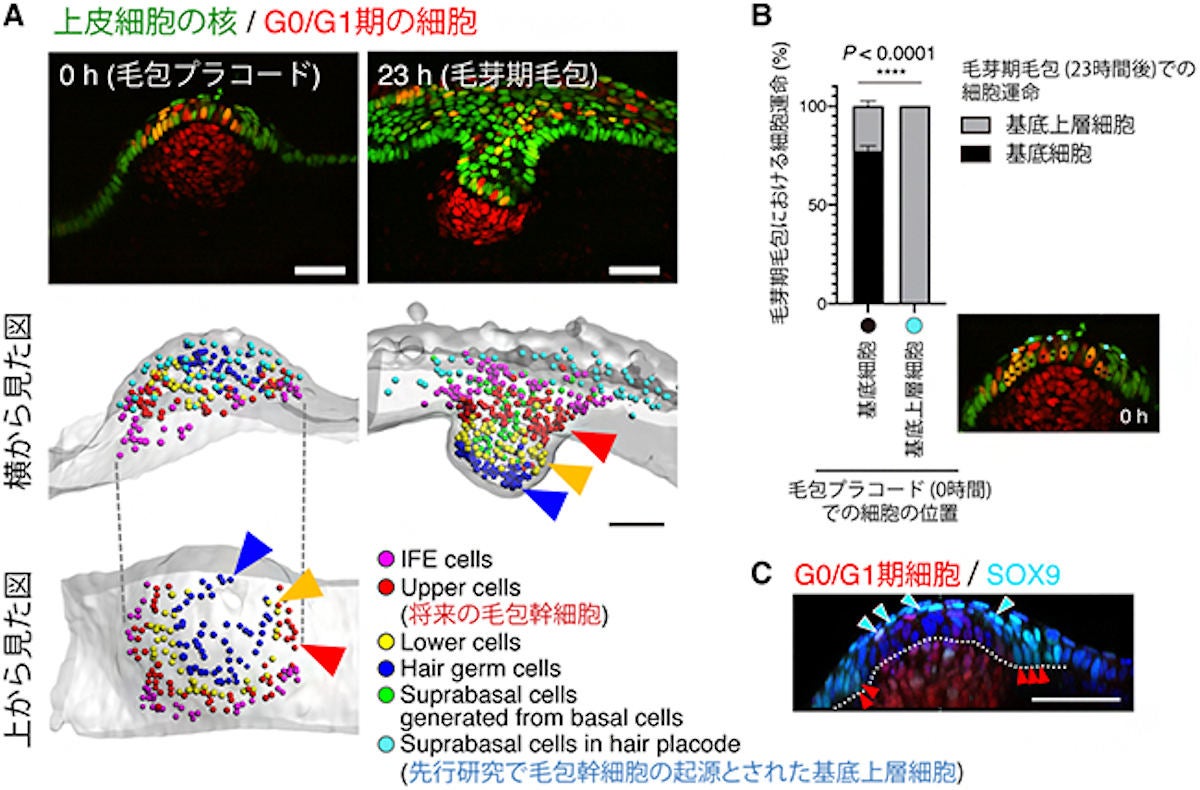
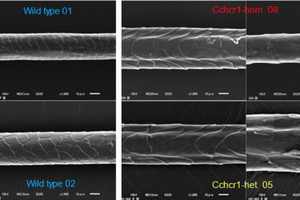
毛包幹細胞の発生予定領域の同定。(A)(上)胎齢12日マウスのプラコード期から毛芽期に至る毛包発生のライブイメージング。(下)毛芽期の各領域の細胞の動きを遡って追跡することで、プラコード期でのそれぞれの発生予定領域が確認可能。毛包プラコードには異なる上皮細胞系譜が同心円上に配置しており、将来幹細胞になる細胞は毛包プラコードの辺縁の基底層(赤矢頭)に存在することがわかった。先行研究において幹細胞の起源とされた基底上層細胞(水色丸)は、毛芽期では表皮内に散在しており、毛包発生にほとんど寄与しないことが判明した。スケールバーは50μm。(B)プラコード期の基底層および基底上層に存在する細胞が、毛芽期毛包ではどこに存在するのかが集計された。基底層を完全に離れた基底上層細胞(水色丸)は、毛芽期において二度と基底層(成体幹細胞が存在する細胞層)に戻らないことが確認された。(C)毛包プラコードには、転写因子SOX9の発現の強さの異なる2種類の細胞が存在していた。先行研究で幹細胞の起源とされた基底上層細胞(水色矢頭)はSOX9を高発現させ、今回の研究で明らかになった幹細胞予定領域の基底細胞は、SOX9を中程度発現させていることがわかった(赤矢頭)。スケールバーは50μm (出所:理研Webサイト)

従来の定説では、基底細胞の非対称分裂により生じたSOX9陽性の基底上層細胞が毛包幹細胞の起源とされていたが、この細胞は分化細胞を生み出すだけで、幹細胞領域には寄与していないことも判明したという。
発生過程での毛包細胞の状態変化を知るための経時的な1細胞トランスクリプトーム解析の結果、毛包プラコードにおいて、ライブイメージングで得られた細胞系譜パターンと一致した同心円リング状の遺伝子発現パターンが存在することが判明したほか、発生ステージの進行とともに、同心円の各リングに特徴的な複数の遺伝子が、同心円パターンから筒状毛包の各区画の発展と対応するように、その発現領域を変化させることも判明したという。
-
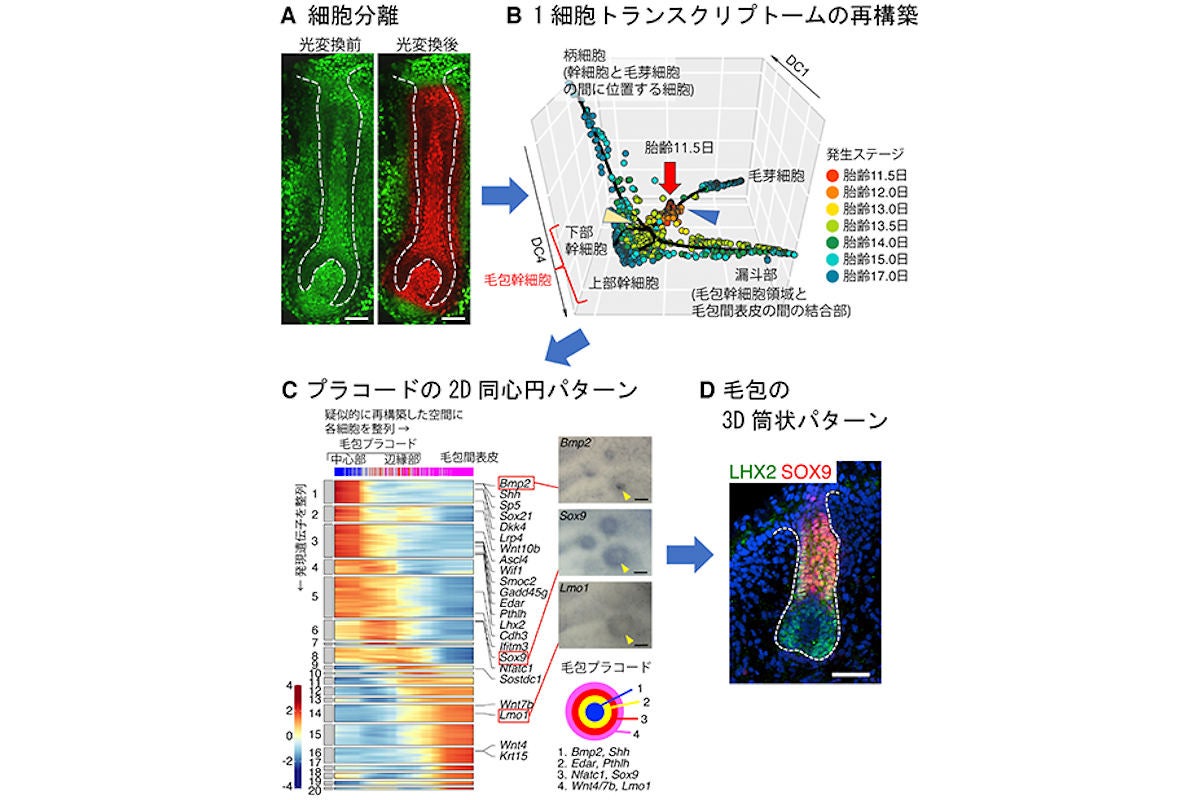
毛包上皮細胞の網羅的1細胞トランスクリプトーム解析。(A)光変換型蛍光タンパク質KikGRを用いて標識された毛包。スケールバーは50μm。(B)発生期の毛包上皮細胞の網羅的な疑似時系列解析。各発生ステージの毛包上皮細胞の1細胞トランスクリプトームデータが用いられ、個々の細胞の遺伝子発現の特徴が三次元空間上の位置で表現されたもの。似た特徴を持つ細胞は同じ位置にクラスターを作る。胎齢11.5日プレプラコード期(赤丸)を起点に、胎齢12日プラコード期(オレンジ丸)を経て、徐々に細胞運命が分岐して、それぞれの細胞系譜ごとに分化・成熟していく様子が捉えられている。青矢頭は毛芽細胞の分岐点が、黄矢頭は毛包幹細胞の分岐点が示されている。(C)胎齢12.0日毛包上皮細胞由来の1細胞トランスクリプトームの詳細な解析。(左)遺伝子発現パターンから擬似的に再構築された空間に1細胞トランスクリプトームデータが並べられた結果、毛包プラコードの中心部から辺縁部、毛包間表皮細胞に向かって発現遺伝子が変化していく様子が認められた。(右上)擬似的に再構築された空間で、プラコードの中心から辺縁に向かって異なる発現パターンが示された3つの遺伝子について、実際の毛包プラコード上での発現パターンが確認されたところ(矢頭)、Bmp2の発現を中心にリング状の発現パターンが観察された。(右下)毛包プラコードで発現する代表的な遺伝子の発現パターンがまとめられたもの。スケールバーは100μm。(D)毛包プラコード上で、リング状の発現パターンを示した転写因子SOX9は、発生が進んだ毛包では円筒状の発現パターンを示すようになる。スケールバーは50μm (出所:理研Webサイト)

研究チームは、この伸縮式の望遠鏡が伸びるように発生する毛包の形態形成様式に対して「テレスコープモデル」と命名。これは、ショウジョウバエ胚発生における肢原基の形成機構にもよく似ていることから、哺乳類以外の生物種の器官発生にも共通する普遍的な形態形成システムである可能性があるという。
-
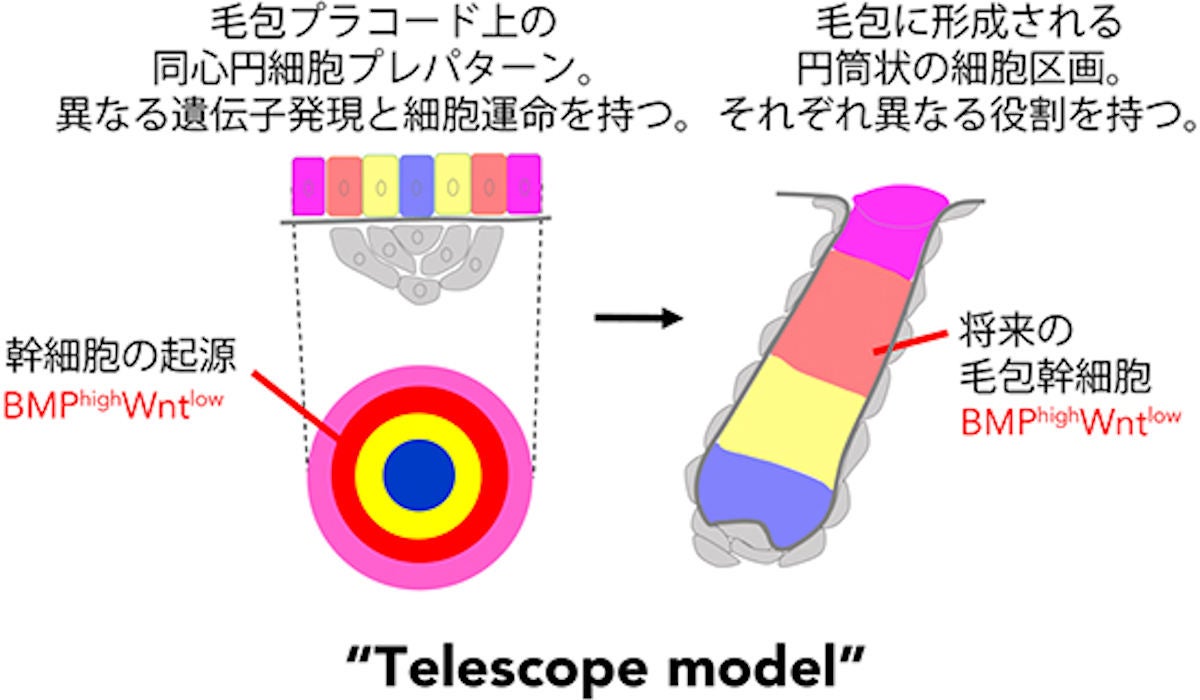
毛包幹細胞の発生起源と新たな毛包発生モデル「テレスコープモデル」。毛包プラコードには、異なる遺伝子発現と細胞運命を持つ上皮細胞が同心円状に配置された毛包のプレパターンがすでに存在している。このうち、毛包プラコード辺縁のBMP活性が高くWnt活性が低い区画から将来の毛包幹細胞が誘導される。このプラコード上の同心円パターンは発生とともに体内側に陥入し、各領域が長軸方向に伸長することで、三次元的な筒状の区画が形成され、毛包構造が作られることがわかった (出所:理研Webサイト)

なお、今回の成果は基礎および応用の両面で発展が期待できるという。基礎面では、幹細胞を生み出し維持する幹細胞の微小環境「幹細胞ニッチ」が解明されると考えられるとしている。また、テレスコープモデルが、毛包と同じようにプラコードから発生する他の体表器官(乳腺や汗腺など)の発生や幹細胞誘導機構にも共通する仕組みかどうかを調べることで、器官や生物種を貫く普遍的な形態形成システムに迫れる可能性があるとしている。
一方の応用面では、ES細胞やiPS細胞など多能性幹細胞からの毛包幹細胞の誘導・増殖・分化の制御技術の発展が期待できるとしており、今回の成果を活用することで、ES/iPS細胞から効率良く質の高い毛包幹細胞を分化誘導できる可能性が高まるとしている。