早稲田大学(早大)と理化学研究所(理研)の両者は7月19日、進化のミッシングリンクとなる新しいタンパク質構造を発見し、これを用いることで、遺伝子発現系に重要なタンパク質構造の進化を実験的に再現することに成功したと共同で発表した。
同成果は、早大 人間科学学術院の八木創太講師(理研 生命機能科学研究センター(BDR) 客員研究員兼任)、BDRの田上俊輔チームリーダーらの共同研究チームによるもの。詳細は、英オンライン科学誌「Nature Communications」に掲載された。
細胞内の遺伝子発現機構では、DNAから転写反応によりRNAが作られ、RNAの翻訳反応によってタンパク質が作り出される。これまで、同機構の詳細な制御の仕組みについて数多くの研究が行われてきたが、太古の地球上で同機構がどのようにして成立してきたのかについては、まだ多くの謎が残されているという。
転写反応ではタンパク質の一種の「RNAポリメラーゼ」が、翻訳反応では、RNA・タンパク質複合体の「リボソーム」がそれぞれ重要な役割を担う。それらは巨大な構造を持つが、初期の生命が最初からそのような複雑な分子を持っていたとは考えにくく、当時はもっと小さくて単純な分子だったと推定されている。
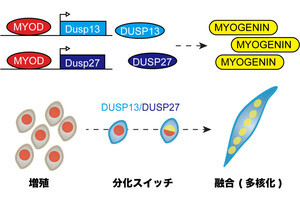
両者の共通項は、小型の「βバレル型構造」を重要な構成要素として有している点だ。たとえば、RNAポリメラーゼの活性コアを形成する「Double-psiβ-barrel(DPBB)構造」、リボソーム「L3」が持つ「RIFT構造」、複数のリボソームに高度に保存される「OBフォールド」や「SH3フォールド」などが挙げられる。これらの構造は部分的な類似点も多いことから、その進化的関係性についての議論がなされていたが、これらのβバレル構造間のアミノ酸配列の違いは大きく、実際どのようにして多様なβバレル構造が進化してきたのかは未解明だったとする。
-

RNAポリメラーゼとリボソームを構成する4つのβバレル構造と提唱される進化的関係性(出所:早大Webサイト)

そうした中で早大の八木講師らは、DPBB構造が7種類のアミノ酸を43個をつなげた単純なペプチドで再構成できることを実証。この単純なペプチドは水溶液中では特定の構造を形成できないが、マロン酸やリンゴ酸などの有機酸を含む溶液中では結晶化してDPBB構造を作ることがわかっていた。そこで研究チームは今回、この単純なペプチドが硫酸イオンを含む条件でも結晶を形成するのか調べしたという。
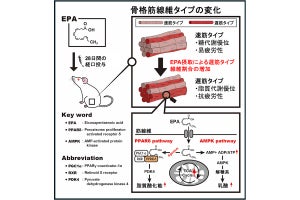
すると、単純なペプチドが硫酸イオンを含む条件でも結晶を形成し、DPBBとは異なるβバレル構造を作ることが見出された。この構造は、これまで自然界では未発見の新たな構造であり、立体構造を模式的に表すとギリシャ文字のZに似た特徴的なループ構造を2つ持つことから、「Double-zetaβ-barrel(DZBB)構造」と命名された。つまり、この単純なペプチドは同じ配列でも異なる条件では異なるタンパク質構造を作るメタモルフィック(変性作用のある)タンパク質であることが判明したのである。
-

単純なペプチドによるDPBB型とDZBB型の結晶構造とトポロジー図。両者で特に異なるループ構造がピンクでハイライトされている(出所:早大Webサイト)

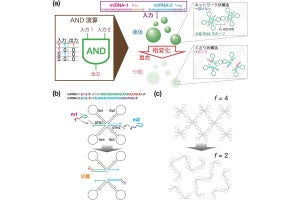
またDZBB構造は、リボソームに含まれるRIFT構造・OB構造と構造類似性が認められたという。そこで、DZBB型タンパク質からRIFT型構造・OB構造への変換が実験的に試みられた。その結果、DZBB-RIFT構造間の変換は1つの短い配列の欠失、DZBB-OB構造間の変換は1つの短い配列挿入と5つのアミノ酸置換のみで達成しうることが突き止められた。さらに、作成されたOB型のタンパク質改変体を循環置換させたタンパク質が、SH3構造を形成することも実証したとする。
研究チームによるとこれらのタンパク質進化の実験は、DPBB、RIFT、OB、SH3の4つの異なるβバレルタンパク質構造が、同じ祖先タンパク質から進化してきたこと、その進化の過程ではDZBBという、今は失われた進化中間体構造があった可能性を強く示唆するという。そしてこの結果は、転写反応を司るRNAポリメラーゼと翻訳反応を司るリボソームが共通の祖先タンパク質から誕生したことを裏付ける証拠だといえるとした。
-

実験的に再現されたDZBB構造から他のβバレル構造への進化の模式図(出所:早大Webサイト)

加えて、今回の研究では従来考えられてきたよりも少ない遺伝子変異だけでタンパク質構造の進化が起こることも発見。つまり、非常に短時間でこれら4種のβバレル構造が誕生した可能性が示されているとしている。
今回の研究成果は今後、リボソームやRNAポリメラーゼなどの巨大な分子がどのように誕生してきたのか、遺伝子発現機構がどのように誕生してきたのかといった謎を探る上で、重要な道標になることが期待できるという。また、タンパク質の初期進化を探求する上でも、重要な視点を与えるものといえるとする。
また研究チームは今後、各βバレルタンパク質がDNAやRNAとどのように結合し、どのように機能するのかを解明していくことで、タンパク質と核酸による共進化の歴史も解明できる可能性があるとしている。