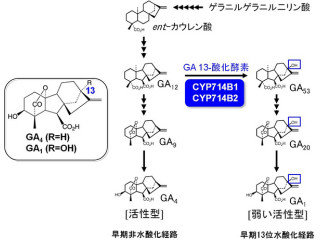
理化学研究所(理研)と東京大学は6月15日、植物の脱分化・器官再生・発生において、遺伝子発現に必須な過程の1つであるメッセンジャーRNA(mRNA)から「イントロン」(遺伝子領域の内タンパク質配列を指定しない領域)を除去する「pre-mRNAスプライシング」と呼ばれるRNA制御機構が重要な働きを持つことを明らかにしたと発表した。
成果は、理研 環境資源科学研究センター バイオマス工学連携部門 セルロース生産研究チームの大谷美沙都研究員、同・出村拓チームリーダー、東大大学院 理学系研究科の杉山宗隆准教授らの研究チームによるもの。研究の詳細な内容は、日本時間6月15日付けで米国植物科学会発行の植物科学専門誌「The Plant Cell」オンライン版に掲載された。
動植物の受精卵細胞のように、1つの細胞が体を形成するさまざまな種類の細胞すべてに分化できる能力を持つことを「分化全能性」という。また、植物細胞は、適切な条件で培養すれば再び根や茎に分化し完全な1つの植物体へと再生できるため、一般的にどの植物細胞も「分化全能性」を保持していると考えられている。これは、胚発生の段階で分化全能性をどんどん失って専門化していく動物細胞と対称的だ。ただし、どのような分子機構によるものなのかについてはよくわかっていないし、さらに植物の正常な発生制御とどう関連しているのかという知見もまだまだ限られている状態である。
近年の環境問題の深刻化は、「カーボンニュートラル」(二酸化炭素の排出と吸収の差がゼロになること)の理念に則った新たな産業形態の創出が求められているが、そのためには有用植物の効率のよい増殖生産システムの構築が欠かせない。有用植物の分子育種の点からも、どのような分子メカニズムが再生能力と関連しているのかを明らかにすることが求められている。そこで研究チームは、植物細胞が持つ高い再生能力を支えている分子機構の解明を目指した次第だ。
研究チームはモデル植物のシロイヌナズナを用いて、温度感受性突然変異体「rid1-1(root initiation defective1-1)」を材料に分子遺伝学的解析を進めた。当初、この変異体は胚軸(発芽した幼植物体の茎部分)からの根の再生に関する変異体として単離されたが、解析によってrid1-1変異体では胚軸脱分化(カルス形成)過程(画像1)のほか、側根形成や「シュート」(茎とそれについている葉をまとめた呼び方)再生の過程も高温になると阻害されることがわかった。こうした表現型は、原因となる遺伝子であるRID1が、脱分化および器官再生において重要な役割を担っていることを意味している(画像2)。
画像1は、rid1-1変異体では温度依存的に胚軸からのカルス形成が阻害されている様子を野生型と比較撮影したもの。野生型(正常)およびrid1-1変異体の胚軸断片をカルス誘導培地上で22℃あるいは28℃で培養した。rid1-1では28℃で培養した場合にカルス形成が起こらない。
そして画像2は、脱分化・器官再生に関するrid1-1変異体の温度感受性をまとめた模式図。rid1-1変異体で異常になる過程が赤矢印で示されている。rid1-1変異体では胚軸からのカルス形成、シュート再生、および根の正常な発達が強く阻害されるが、こうした表現型からRID1遺伝子は脱分化および器官再生に必要であると考えられた。

|

|
|
画像1。野生型とrid1-1変異体の温度依存的に胚軸からのカルス形成の差 |
画像2。脱分化・器官再生に関するrid1-1変異体の温度感受性をまとめた模式図 |
RID1遺伝子について詳しく調べると、タンパク質「DEAH-box型RNAヘリカーゼ」をコードしていること、rid1-1変異体ではこの遺伝子配列の内の1塩基が置換されることで、RID1タンパク質を構成するアミノ酸の1つが別のアミノ酸に置換されていることが判明。このRNAヘリカーゼは、酵母やヒトで遺伝子からタンパク質が合成される過程で重要な「pre-mRNAスプライシング」に関連するタンパク質によく似ていた。そのため、RID1もpre-mRNAスプライシングに関与している可能性が考えられた。
そこで、rid1-1変異体と野生型の細胞にタンパク質をコードしないイントロンを含む遺伝子を導入してスプライシング効率を比較したところ、rid1-1変異体の細胞でスプライシング効率が著しく低下していることがわかったのである(画像3・4)。
なお、pre-mRNAスプライシングについて捕捉すると、遺伝子発現のためには、最初に遺伝子がタンパク質翻訳の鋳型であるmRNAへと転写され、その転写直後のmRNAはpre-mRNA(プレmRNA)と呼ばれる前駆体の状態だ。真核生物の場合、多くの遺伝子はタンパク質配列を指定する領域の「エクソン」とそうでないイントロン両方を含んでいるが、この内イントロンが最終的にはmRNAから取り除かれる必要がある。このイントロン除去過程をpre-mRNAスプライシングと呼ぶ。pre-mRNAスプライシングは核内で起こり、イントロンが除去された成熟mRNAは核外へ運ばれ、タンパク質に翻訳されるという仕組みだ。
画像3は、実験に用いた人工作製したレポーター遺伝子の模式図。人工的に遺伝子内にイントロンが挿入されている。これを細胞内に導入することでpre-mRNAスプライシングが正常に起こると短い断片が、起こらないと長い断片がそれぞれ検出される仕組みだ。レポーター遺伝子とは、遺伝子組換えが行われた時に導入した遺伝子が発現しているかどうか確認するために用いられる遺伝子のことである。
画像4は、実験の結果。野生型ではスプライシングが起こり短い断片が優勢となるが、rid1-1変異体ではスプライシングされていない長い断片がより多く検出された。また、この長い断片は28℃の時により顕著に蓄積していた。

|

|
|
rid1-1変異体では温度依存的にpre-mRNAスプライシング効率が低下する。画像3(左):実験に用いられた人工作製したレポーター遺伝子の模式図。画像4:実験結果 |
|
また、rid1-1変異体では「選択的スプライシング」が起こることが知られている遺伝子の発現にも野生型とは異なるパターンが見られ、変異体ではmRNA上のどの領域をイントロンとして認識するかが乱れていることがわかった(画像5)。
選択的スプライシングとは、1つのpre-mRNAから複数タイプの成熟mRNAが作られる場合のことをいう。これによって、エクソン-イントロン構造などが変化したmRNAが生み出され、1つの遺伝子から複数のタンパク質が作られることもある。
rid1-1変異体では温度依存的に選択的スプライシングパターンが変化することが確かめられた結果をまとめたものが、画像5だ。選択的スプライシングが起こることが知られているAt4g25500遺伝子についての発現が調べられたところ、rid1-1変異体では赤い四角で囲ったタイプのmRNAが減少していることがわかった。右側の図の黒い四角はエクソンとして認識された領域を、黒線はイントロンとして認識されて除去された領域を示している。

|
|
画像5。rid1-1変異体では温度依存的に選択的スプライシングパターンが変化することが確かめられた |
以上から、RID1はpre-mRNAスプライシング制御因子であることが示されたというわけだ。RID1は、pre-mRNAスプライシング進行の場である核質ではなく、核小体と呼ばれる核内小器官に局在している(画像6~8)。核小体はpre-mRNAスプライシングを担うsnRNPと呼ばれるRNA-タンパク質複合体形成の場であることから、RID1もsnRNP形成に関与していると考えられるという。

|

|

|
|
RID1タンパク質は主に核小体に局在する。画像6(左):RID1-YFPシグナル。RID1タンパク質に蛍光タンパク質YFPを融合し、根の細胞で発現させたもの。緑色に見えるのがシグナル。画像7(中):DAPI染色シグナル。画像6と同じ細胞を蛍光染色試薬「DAPI」で染色した細胞像。DAPIは核酸と優先的に結合する試薬で、赤紫色に見えているのがシグナルだ。細胞中心の丸く見えている構造が核で、赤紫色が強い部分が核質、黒く抜けているように見える部分が核小体である。画像8(右):画像6と7を重ね合わせた像。緑色のシグナルが核小体に存在しているのがわかる |
||
また研究チームは、RID1機能不全が植物発生にどういう影響をもたらすのかの詳細な解析も実施した。rid1-1は28℃で育てると、芽生えの段階で致死となる。この時、新たな器官を生み出す頂端分裂組織が正常に確立・維持されず、結果的に死に至っていることがわかった。
また、生存可能な22℃で育てた場合でも、rid1-1では葉での維管束形成など細胞分化が異常になっていた。さらにRID1遺伝子の機能が完全に失われるノックアウト変異の解析も実施され、めしべで作られる卵細胞などの形成が異常となっていることが判明。RID1遺伝子は、こうした変異体で異常が現れる器官や組織で強く発現していたのである。これらから、RID1が持つ機能、すなわちpre-mRNAスプライシング制御は、分裂組織形成や配偶体形成など、特定の植物発生制御で重要であることが示唆されたというわけだ(画像9)。
画像9は、RID1機能が必要とされる植物の発生過程のまとめたもので、rid1変異体で異常となる過程が赤字で示されている。RID1機能はこれらの発生過程で特に必要とされる。写真は28℃における野生型とrid1-1変異体の幼植物体の12日目における様子だ。rid1-1は野生型と異なり新しい葉が形成されないし(左)、側根の形態も異常になってこぶ状になる(右)。

|
|
画像9。RID1機能が必要とされる植物の発生過程のまとめ |
pre-mRNAスプライシング制御異常がなぜ細胞分化・増殖の阻害を引き起こすのかが今回はわからなかったが、研究チームはその詳細なメカニズムの解明にすでに取り組んでいるとした。その阻害の原因となっている分子が明らかになれば、植物細胞の分化や増殖を人為的にコントロールする技術開発につながると考えられるという。これは、効率的なクローン増殖が難しい早生樹などの有用植物種に対して、有効な新規バイオテクノロジーとなることが期待できるとしている。