理化学研究所(理研)は1月15日、植物生長ホルモンの1つで、多くの植物に普遍的にある「ジベレリンA1(GA1)」の生合成に関わる酵素遺伝子の中で、唯一未解明だった「GA13-酸化酵素遺伝子」をイネから発見したと発表した。
成果は、理研 植物科学研究センター 生長制御研究グループの山口信次郎客員主管研究員(現・東北大学大学院生命科学研究科教授)、同・真籠洋研究員、同・神谷勇治グループディレクター、宇都宮大学の野村崇人准教授、静岡大学の大西利幸特任助教、東京農工大学大学院連合農学研究科の川出洋准教授らの研究グループによるもの。研究の詳細な内容は、1月14日付けで米国科学雑誌「米科学アカデミー紀要(PNAS)」に掲載された。
植物ホルモンは植物自身が合成し、シグナルとして自身の細胞にさまざまな働きかけをする生理活性物質だ。一般に植物組織1g中の植物ホルモンの含量は、1ng以下と極微量で、その量は生長や環境に応じて巧妙に調節されている。
植物ホルモンの1つであるジベレリン(GA)は、植物の生長を促す作用を持つ。1940~60年代にコムギやイネの収量を飛躍的に高めた「緑の革命」では、草丈がやや低い品種の特性を人為的な交配によって取り入れて、肥料を多く与えても倒れにくい多収品種の開発につながった。この草丈が低い特性は、GAの生合成遺伝子などの突然変異が原因であることが、後の研究で明らかにされている。
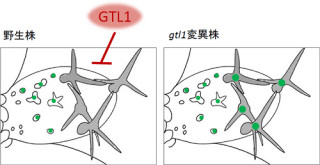
GAは、多段階の酵素反応を経て生合成される仕組みだ。合成中間体であるGA12以降は、GA13-酸化酵素遺伝子による13位水酸化(OH基の導入)への変換の有無により、早期非水酸化経路と早期13位水酸化経路に分かれる。そして、その後も既知の酵素により順次酸化され、最終的に活性の強いGA4(13位非水酸化型)と活性の弱いGA1(13位水酸化型)が合成される(画像1)。

|
|
画像1。イネのジベレリン生合成経路。GA13-酸化酵素が、GA1(弱い活性型)の生合成経路のカギとなる。イネの茎葉ではGA1が主要な活性型である |
GA1は、イネを含む多くの種子植物で普遍的に見られる活性型のGAだ。そしてGA13-酸化酵素は、GA1の生合成に欠かせないカギとなる酵素である。以前からその酵素活性はいくつかの植物で見つかっていたが、それをコードする遺伝子は長らく不明だった。
これまでに、GAの生合成に関わるほかのすべての酵素遺伝子は明らかになっていたため、ただ1つ未解明なGA13-酸化酵素遺伝子はGA生合成のミッシングリンク(発見されていない間隙)といわれていのである。
研究グループは、2006年に中国のグループとの共同研究でイネから発見した、GAを不活性化させる「シトクロムP450酵素」をコードする「EUI(CYP714D1)遺伝子」に着目。
この遺伝子に遺伝子配列が近いイネゲノム上の5つのCYP714ファミリーに着目し、詳しく調べた。これらを、モデル植物であるシロイヌナズナで過剰発現させた結果、CYP714B1とCYP714B2遺伝子の各過剰発現体が野生型に比べて小さくなった。そして、体内のGA含量を分析した結果、野生型に比べGA1(13位水酸化型)が増えており、13位水酸化が亢進していた。
次に、これらがコードする酵素とGAとの関連性を直接検証するため、酵母や昆虫細胞でCYP714B1酵素とCYP714B2酵素を各々発現させる形で、機能解析が行われた。その結果、両酵素にGA中間体であるGA12の13位を水酸化しGA53に変換させる活性があることを明らかにした。この結果は、GA12が(GA1が作られる)早期13位水酸化経路へとつながることを意味する(画像1)。
さらに、2つの遺伝子を欠損させたイネ二重変異体のGA含量を分析したところ、GA1が著しく減少した代わりにGA4が蓄積していた(画像2)。同時に測定したほかのGA中間体の測定結果から、このGA1の減少とGA4の蓄積は、13位水酸化が抑制された結果によるものと判明したのである(画像2)。

|
|
画像2。cyp714b1cyp714b2の二重変異体のGA量の分析結果。*印は検出限界以下。二重変異体では、活性型GA1を含む早期13位水酸化経路のGAが低下しており、反対に活性型GA4を含む早期非水酸化経路のGAが増加していた |
これらの結果から、CYP714B1とCYP714B2はイネの主要なGA13-酸化酵素遺伝子と判明した。また、この二重変異体は、出穂期に最上位の節間が野生型と比べて長くなり草丈が高くなった(画像3)。

|
|
画像3。野生型と変異体の様子。一重変異体間の交配により得られた雑種第2(F2)世代。左より分離野生型、cyp714b1変異体、cyp714b2変異体、cyp714b1cyp714b2二重変異体(バーは20cm)。二重変異体は最上位の節間が長いため、草丈が高くなる |
これらから、イネは時期と組織に応じてGA4とGA1の量を調節して生長を最適化していることがわかった。イネの茎葉にはGA1が多く存在する一方、GA4は葯に非常に多く存在することが知られている。また、GA1はGA4に比べ活性が低いだが、前述のEUI(CYP714D1)酵素による不活性化を受けない。これまでGAの活性は、その生合成と不活性化酵素の巧妙なバランスによる量の調節が働いていることが知られていた。今回の研究によりその量の調節に加え、GA13-酸化酵素の働きによる質の調節、すなわちGA1とGA4の量比の調節も働いてGAの活性が制御されることが示唆された。
今回の成果により、GA生合成調節の全体像の詳細な解明が進み、その先の「なぜ植物は複数の活性型GAを使うのか」という大きな疑問の答えが得られるのではないかと期待している。
また、シロイヌナズナやキュウリなどの茎葉ではGA4が多く存在している。今回のイネのGA13-酸化酵素遺伝子を過剰発現させたシロイヌナズナが野生型に比べ小さくなったのは、GA4とGA1の量が逆転したために起きたと考えられる。草丈の適度な抑制は農作物の生産性向上につながる可能性を持っている。このGA13-酸化酵素を用いたGAの質の調節による、新しい植物生長調節技術の開発が期待できる。