科学技術振興機構(JST)と名古屋大学(名大)は5月14日、被子植物独自の生殖システムである卵細胞と「中央細胞」の2つに受精する「重複受精」を人為的に制御し、それぞれの細胞に異なる個体の精細胞を受精させる「ヘテロ受精」をシロイヌナズナで実現すると同時に、花粉の精細胞を雌しべの奥の胚のう内にある卵細胞と中央細胞に届けるために伸長していく「花粉管」の誘引を停止する仕組みも発見したと共同で発表した。
成果は、名大 WPI トランスフォーマティブ生命分子研究所の丸山大輔研究員、JST戦略的創造研究推進事業ERATO型研究「東山ライブホロニクスプロジェクト」の東山哲也研究総括(名大 WPI トランスフォーマティブ研究所 教授)らの研究チームによるもの。研究はJST課題解決型基礎研究の一環として行われ、詳細な内容は、米国東部時間5月13日付けで米科学誌「Developmental Cell」オンライン速報版に掲載された。
重複受精は、被子植物の最もユニークな特徴の1つだろう。卵細胞と中央細胞という2つの生殖細胞があり、それぞれを独立して受精させ、「胚(種子の中で植物体になる部分)」と「胚乳(種子の中の栄養器官)」を作る仕組みである(画像1)。よって、植物の卵細胞と中央細胞を、異なる遺伝子を持つ精細胞と受精させることができれば、胚と胚乳に異なる遺伝情報を持たせるよう制御することができるというわけだ。

|
|
画像1。被子植物は、重複受精という仕組みで、種子の内部に胚と胚乳を形成する |
植物の受精というと、「雌しべに花粉がつくこと」というイメージが一般的だ。別にそれが間違いではないが、厳密にはそこで受精そのものが完了するわけではない。雌しべ深部にある「胚のう」(1個の卵細胞、1個の中央細胞、2個の「助細胞」、3個の「反足細胞」で構成される、受精後に種子を形作る中心組織)内の助細胞からの誘引を受けて、花粉からその雌しべ深部へと長い管上の細胞である花粉管が伸びていく。被子植物の精細胞は鞭毛を欠いていて自ら泳ぐことが不可能なことから、花粉管の先端内部にある2個の精細胞は、花粉管自体が卵細胞まで伸びていって到達することで初めて受精が起き、種子が形成されるというわけだ。
1本の花粉管によって同じ遺伝子組成の2つの精細胞が運ばれ、それらが卵細胞、中央細胞に届くことで受精が完了する。そして受精が完了すると、花粉がどれだけ雌しべについていようと、それらが花粉管を伸ばしている途中であろうと、助細胞による誘引は終了となる形だ。よって、胚と胚乳に異なる遺伝情報を持たせるような制御は難しいと考えられてきた。
しかしトウモロコシは別のようで、まれに胚と胚乳が異なる遺伝子を持つ精細胞と受精する「ヘテロ受精」が自然に起きることが、1929年に報告されている。しかしそれから長い年月が経過したにもかかわらず、これまでのところ、どのようにヘテロ受精が起きるのか、その仕組みは解明されてこなかった。そのこともあって、胚と胚乳を異なる遺伝子組成にして両者を制御することは原理的には可能であるものの、人為的な制御は難しいと考えられるようになっていたのである。
そうした状況の中、研究チームは今回、重複受精における2つの雌しべ側の生殖細胞(卵細胞と中央細胞)の受精と、花粉管の誘引停止の関係を明らかにするために、アブラナ科植物であり、モデル植物としてよく利用されるシロイヌナズナを用いて、従来にない研究に挑んだ。
被子植物の重複受精は、雌しべの奥深くでほぼ同時に起きるために、卵細胞と中央細胞の受精を区別して解析することが困難だ。そこで、通常2つある精細胞が1つしか作られない突然変異のあるシロイヌナズナ「cdka;1」変異体に白羽の矢が立った。
この変異体の花粉を授粉させると、卵細胞と中央細胞のどちらか一方との受精が起こるため、胚または胚乳のどちらかのみが発達した種子が作られる。研究チームがこれらの種子を調べたところ、どちらのタイプでも花粉管の誘引は完全には停止せず、約30%の割合で複数の花粉管を誘引していることが判明した。こうした解析から、以下の2点が関係性があると推論されたのである。
(1)卵細胞と中央細胞は、それぞれ受精することで2本目の花粉管の誘引を止めるように作用するが、どちらか一方の受精だけでは不十分で、花粉管の誘引を完全に止めることができない。(2)卵細胞と中央細胞が両方とも受精すると、協調的に作用し2本目の花粉管誘引を停止できる(画像2・3)。
また、卵細胞と中央細胞での花粉管誘引の制御は独立して起こり、中央細胞では胚乳形成を制御する遺伝子としてよく知られていた「MEDEA遺伝子」が、花粉管の誘引停止に必要であることもわかった。

|

|
|
卵細胞と中央細胞による花粉管の誘引停止の模式図。画像2(左):卵細胞と中央細胞の受精がそれぞれ独立に、花粉管を誘引する助細胞に作用し、花粉管の誘引が完全に停止する。画像3(右):片方の受精(単独受精)でも、誘引の低下が見られるものの、花粉管誘引は完全には停止しない(左)。このため、2本目の花粉管が誘引され、ヘテロ受精が起こる(右) |
|
これらの結果から、今度は卵細胞または中央細胞の片方だけが受精に成功した雌しべ側の生殖細胞について、次に誘引された花粉管で重複受精が完了する、すなわちヘテロ受精が起こるのではないかと研究チームは推論。さらに、解析が進められたのである。
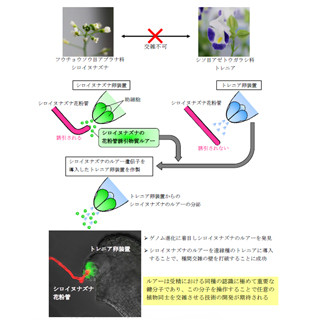
次の実験には、2つの精細胞の内の片方だけが受精する傾向を示す突然変異体のシロイヌナズナ「kokopelli」変異体が用いられた。胚と胚乳の細胞核の色が異なる種子が観察できるようにするため、kokopelli変異体を改変して、2種類の変異体を用意。最初は、細胞核が顕微鏡下で赤色に光るようにした変異体の花粉を用いて、正常なシロイヌナズナへの授粉が行われた。そしてその3時間後には、今度は細胞核が顕微鏡下で緑色に光るように改変された変異体の花粉による授粉が行われたというわけだ。
これらの2種類の変異体を利用して時間差をつけて花粉を授粉させることにより、初めのkokopelli変異体の授粉で卵細胞または中央細胞のいずれかと受精し、次の授粉で残った方の受精が起きるというわけである。
実験の結果、胚と胚乳の細胞核の色が異なる種子を得ることに成功(画像4・5)。シロイヌナズナでヘテロ受精が示されたのは初めてのことだ。また、ヘテロ受精を人為的に引き起こすことができることも明らかとなった。これまで謎だったヘテロ受精には、卵細胞と中央細胞のそれぞれの受精による協調的な花粉管誘引停止の仕組みが背景にあることが判明したのである。

|

|
|
ヘテロ受精の人為的な制御。画像4(B):重複受精は同じ遺伝子組成を持つ1対の精細胞により達成される。そのため、緑色に光るタンパク質を発現するような精細胞で受精が起こると、胚も胚乳も緑色に光る。画像5(C):kokopelli 変異体を用いて人為的にヘテロ受精を起こすことを試みた結果、緑色に光るタンパク質を発現する精細胞と、赤色に光るタンパク質を発現する精細胞で受精が起こり、胚と胚乳が異なる色で光る種子が得られた。それぞれの下段では、上段と同じ組織における花粉管(PT)を可視化している。画像5では花粉管が2本到達していることが確認できる |
|
研究チームは、今回の成果をもとにすることで、卵細胞と中央細胞のそれぞれがどのように協調的に花粉管誘引を停止するのか、その仕組みの解明が期待されるという。今回の成果は、ドイツのグロスハート博士らの論文と連報で誌面を飾る予定だ。グロスハート博士らは、花粉管の誘引停止には、誘引を司る助細胞に「エチレン」が作用して細胞死を引き起こすことが必要であることを示した。卵細胞や中央細胞の受精がどのようにエチレンの生成に関わるのか、今後の研究の展開が期待されるという。
また、ヘテロ受精を人為的に起こすことができるようになったことは、基礎研究と応用研究の両面での展開が期待されるとする。胚と胚乳は種子の中で協調して形成が進行するのはこれまで述べた通りだ。その際の胚と胚乳の間での生成物のやり取り(コミュニケーション)が、両者の発達に対して極めて重要であることが知られている。
しかしこれまでは、それぞれを異なる遺伝子組成にすることが不可能であったために、胚と胚乳で生成される物質の区別をつけることが難しく、どのようなコミュニケーションが行われているのか、具体的には明らかになっていない。今回の研究成果によりヘテロ受精を人為的に制御できるようになったことで、胚と胚乳の相互作用による種子形成の仕組みの解明が期待されるとしている。
さらに応用面では、卵細胞と中央細胞の受精を独立に制御することで、さまざまな展開が期待されるという。例えば、胚と胚乳が異なる発生プログラムで成長するため、それぞれに最適化した遺伝子を持たせることで、よく発達した、貧栄養の土地でも育ちやすい種子を得ることも可能になるとことが期待されるとする。
そのほかにも、育種において異なる植物種を交雑させ新しい植物種を得る際に、胚と胚乳のコミュニケーションがうまくいかないために種子形成ができず、雑種(新しい植物種)が得られないことがあるが、そうした不和を取り除いたり、あるいは雑種の胚の成長をサポートするような胚乳を作り出したりすることで、新しい有用な植物の作出につながる可能性も考えられるという。このように、今回の研究成果でヘテロ受精の謎が解明され、人為的な制御が可能になったことで、植物の生殖制御技術の新しい扉が開いたといえるとした。