東京大学は5月7日、麹菌において「オートファジー(自食作用)」機能を抑制することで、異種タンパク質の生産性が向上することが発見されたと発表した。
成果は、東大大学院 農学生命科学研究科 応用生命工学専攻菊間隆志特任助教、同・Jaewoo Yoon特任助教(当時)、同・丸山潤一助教、同・北本勝ひこ教授らの研究チームによるもの。研究の詳細な内容は、4月29日付けで米オンライン科学誌「PLoS ONE」に掲載された。
大量のタンパク質を培地中に分泌生産する能力を有するのが麹菌の特徴だ。安全性の面に関しても申し分なく、日本においては1000年以上もの昔より発酵食品の醸造に用いられてきている。こうした特徴があることで、麹菌は異種タンパク質生産の宿主としてが非常に優れているというわけだ。しかし、麹菌において高等生物由来の異種タンパク質を発現させると、期待通りの生産量が得られない場合が多い。これは、転写、翻訳、分泌経路などにおいてさまざまなボトルネックが存在するからであると考えられている。
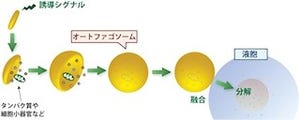
研究チームでは以前、麹菌で変異型分泌タンパク質として正常な「S-S結合」が形成できない「α‐アミラーゼ」を発現させると液胞に取り込まれることを見出し、これがオートファジーに依存することを明らかにした実績を持つ。なお、S-S結合とはジスルフィド結合とも呼ばれ、タンパク質を構成するアミノ酸の1つである「システイン残基」同士でできる硫黄原子(S)を介した結合のことだ。この結合ができないと、タンパク質の正しい折りたたみが行われない。
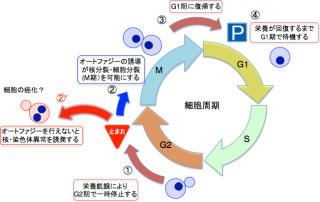
またオートファジーはヒトをはじめとする真核生物に高度に保存された細胞内分解機構であり、栄養飢餓時において細胞質成分を膜で囲い込み、液胞に輸送して分解しリサイクルする生存戦略として機能している。またオートファジーは、発生や分化、免疫応答、細胞死などにも重要な役割を果たしており、さらにがんを初めとするさまざまな疾患に関与していることが知られている重要な仕組みだ。
そして研究チームは今回の研究で、前述した異種タンパク質生産のボトルネックとして、オートファジーこそがその1つであると推測。麹菌の異種タンパク質生産性について、オートファジー欠損株を用いて検討が行われた。
まず、オートファジー関連遺伝子「Aoatg」破壊株を作製し、それぞれの遺伝子破壊株で異種タンパク質のモデルとして「ウシキモシン」を発現させた。ウシキモシンは「レンニン」とも呼ばれ、チーズ製造に用いられるウシの第4胃に存在する凝乳酵素である。その結果、親株と比較して生産量を2~3倍増加させることに成功した(画像1)。

|
|
画像1。Aoatg遺伝子破壊株のウシキモシン生産量。Aoatg遺伝子を破壊するとウシキモシン生産量が増加する |
しかし麹菌のオートファジー欠損株は、通常の培地での増殖は野生株と差はないものの、「分生子」(カビなどが無性生殖の方法として形成する胞子)をほとんど形成しない。このため、これらの株を工業的な規模で異種タンパク質生産に用いる場合、植菌のために必要な分生子を十分に得ることが困難であるという問題が生じる。
そこで研究チームは、これらの遺伝子が分生子形成時には発現し、ウシキモシン生産時は発現しないように制御できる「Aoatg遺伝子発現制御株」を作製。その結果、十分量の分生子を得ることに成功し、またウシキモシン生産量が破壊株と同程度まで増加する実用的異種タンパク質生産株の育種に成功したのである。
分泌タンパク質は、細胞内小器官の1つである「小胞体」で適切な形に折りたたまれることによって機能的なタンパク質となり、分泌経路の次のステップである同じく細胞内小器官の1つである「ゴルジ体」へと輸送される流れだ。しかし、異種タンパク質を多量に発現させるとその折りたたみが間に合わず、凝集体を形成するものが多くなり、そのようなものはオートファジーで分解されると考えられるという。
ところがオートファジーを抑制した場合、凝集タンパク質が分解されないため、小胞体内で折りたたみが何度も行われ適切な形になることにより、多量に発現した異種タンパク質が分泌経路によって菌体外に分泌されると考えられるのである。
画像2は、オートファジー抑制によるウシキモシン生産上昇のモデル図。野生株ではオートファジーにより小胞体で凝集したウシキモシンが分解されるが、オートファジー欠損株ではこの分解が起こらず、分泌経路に入るウシキモシンが増加すると考えられる。

|
|
画像2。オートファジー抑制によるウシキモシン生産上昇のモデル図 |
今回の研究は、糸状菌でオートファジーという基礎生物学的研究と、異種タンパク質生産という応用的研究をリンクさせた初めてのものだという。これらの結果は、麹菌による有用タンパク質の効率的生産にとどまらず、オートファジーの新規機能の発見にもつながるものと期待されると、研究チームは語っている。