理化学研究所(理研)は7月16日、米アルバート・アインシュタイン大学との共同研究により、従来では不可能だった100個程度の細胞からの「DNAメチル化」解析を可能とする技術を開発し、同技術を用いてマウス胎児などから得られる少数の細胞を解析したところ、生殖細胞特有な遺伝子の発現に関わる低メチル化DNA領域が発見されたと発表した。
成果は、理研 バイオリソースセンター 疾患ゲノム動態解析技術開発チームの阿部訓也チームリーダー、同・池田理恵子テクニカルスタッフ(筑波大学大学院博士課程在籍)、同・志浦寛相基礎科学特別研究員、アルバート・アインシュタイン大のスタッフらの国際共同研究チームによるもの。研究の詳細な内容は、7月16日付けで日本の科学雑誌「DNA Research」オンライン版に掲載された。
生物を構成する細胞は、生殖細胞(精子、卵子など)と体細胞に大別される。体細胞は身体を形作るのに必要で個体1代限りの細胞だが、生殖細胞は次代に遺伝情報を受け渡す重要な役割を持っており、世代をつなぐ「不死」の細胞といえよう。
このように体細胞と生殖細胞では異なる役割を持つため、そこで働く遺伝子も当然ながら異なっており、生殖細胞だけで発現し機能する遺伝子が多数存在する。中には極端なものもあり、例えば、雄の生殖細胞特有の遺伝子の一部は、正常な体細胞では発現しないが、がん細胞では発現することがあるのだ。これらの遺伝子は、「がん精巣抗原」遺伝子と呼ばれている。
がん精巣抗原はがん細胞の同定に有効であり、またがんの免疫治療の標的となる可能性も指摘されているところだ。ヒトには260以上のがん精巣抗原遺伝子が存在し、その4割以上が性染色体であるX染色体にあることが知られている。しかし、なぜ生殖細胞とがん細胞で共通して発現するのか、その意義やメカニズムは不明なままだ。
また細胞では、DNAのメチル化や染色体を構成するタンパク質である「ヒストン」(DNAは「クロマチン」という構造を作って核内に収納されているが、その時にDNAが巻き付くタンパク質のこと)のメチル化=「ヒストン修飾」などさまざまな修飾が起きており、細胞ごとに固有の修飾パターンで、遺伝子の発現を制御している。
なおDNAメチル化とは、アデニン・チミン・グアニン・シトシンと4つある塩基の内、ほ乳類の場合に起きるシトシンの炭素原子にメチル基「CH3」が付加される現象のことをいう。このメチル化によりゲノムの構造や機能が変化し、遺伝子発現に影響を与える。関与する生命現象は、発生、細胞分化、ゲノム安定性維持や発がんなどだ。
またヒストン修飾とは、ヒストンにアセチル基やメチル基などが付くことをいう。クロマチンの構造が変わることで遺伝子のオン・オフが調節されるが、その構造変化はDNAメチル化だけではなく、ヒストン修飾によっても制御されている。このように、染色体を構成するDNAやタンパク質が、メチル化や「アセチル化」などの後天的な化学修飾を受けており、この特定の細胞に固有なゲノム修飾状態全体を、「エピゲノム」と呼ぶ。
DNAメチル化は遺伝子発現に関わるため、その制御機構を理解する上で重要だが、生殖細胞で起きるDNAメチル化についてはこれまであまり理解が進んでいなかった。生体内で発生する生殖細胞の数が限られており、通常の手法では解析が困難なことや、生殖細胞と同様な性質を持つ適切なモデル細胞リソースがないことなどがその理由に挙げられる。
そこで研究チームは、生殖細胞に特有な遺伝子発現の制御機構とDNA修飾の関連性を調べることや、生殖細胞の特徴を兼ね備えた細胞リソースを探索することを目的に、マウスを用いて網羅的な遺伝子発現解析とDNAメチル化の解析を実施した。
マウスでは、受精後3.5日目の胚は胚盤胞と呼ばれ、ES細胞(胚性幹細胞)はこの胚から作製される。その後子宮への着床を経て、受精後7.5日の胚で将来の生殖細胞の元となる「始原生殖細胞」が出現。始原生殖細胞からは「EG細胞(胚性生殖細胞)」と呼ばれるES細胞とよく似た多能性幹細胞を作製することが可能だ。始原生殖細胞は体内を遊走して受精後11.5(日)前後に将来の精巣や卵巣の元となる「胎児生殖巣」に到達してその中で配偶子形成が進み、雌雄それぞれの生殖細胞、すなわち精子や卵子が作られるのである(画像1)。

|
|
画像1。マウス生殖細胞系譜と胚体、生殖細胞より作製される幹細胞 |
胎児内に存在する生殖細胞の数はごく限られており、7.5日胚では50個程度、最も多い場合でも1つの胎児から得られる生殖細胞は数1000個にすぎない。そのため、従来のDNAメチル化解析には10万~100万個ほどの細胞が必要なことから、マウス胎児の生殖細胞には適用することが叶わないのである。
そこで研究チームは従来法を見直し、偏りの少ないDNA増幅法など実験条件の最適化や、染色体全域が解析可能なカスタムマイクロアレイの利用などの改良を加えることで、最少100個程度の細胞から得られるDNA(総量が0.5ng以上)から染色体全体のDNAメチル化解析を行える技術を開発したというわけだ。
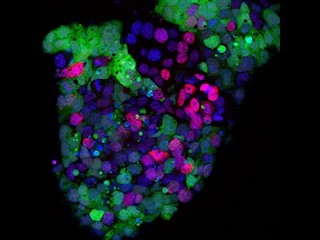
画像2が、今回開発されたDNAメチル化解析技術をまとめたものだ。まず(A)にあるように、生殖細胞で緑色蛍光タンパク質を発現する遺伝子導入マウスを材料に、蛍光発現を目印に生殖細胞を単離する。(上)は胎児精巣内で緑色蛍光タンパク質を発現する生殖細胞、(下)は蛍光発現を指標に単離して純化した生殖細胞だ。
続いて(B)にあるように、ゲノムDNAをメチル化感受性あるいはメチル化非感受性の制限酵素で切断し、それぞれのDNA断片を「リンカーPCR法」で偏りなく増幅させる。メチル化感受性酵素は、非メチル化DNAだけ切断しメチル化DNAは切断されず残るので、増幅されたものは非メチル化DNAに由来することがわかる仕組みだ。一方でメチル化非感受性酵素は、メチル化の有無にかかわらず切断するので、ゲノム全体を含むDNA集団が得られる仕組みである。そして、それぞれを蛍光色素で標識し(例:非メチル=緑、ゲノム全体=ピンク)、ゲノムマイクロアレイと反応させると、低メチル化領域では緑色蛍光の強度が相対的に高くなることからメチル化の程度を判断できるというわけだ。

|
|
画像2。今回開発されたDNAメチル化解析技術。(A):まずマウス胚、胎児からの生殖細胞の単離。(B):微量細胞からのDNAメチル化解析 |
続いて研究チームはこの新しい手法を用いて、マウス胎児から得られる生殖細胞、胚や生殖細胞に由来するさまざまな幹細胞、および数種の体細胞についてDNAメチル化の解析を実施。すると、雄の生殖細胞から低メチル化状態にあるDNA配列が発見された。次にそのDNA配列が染色体上のどこに存在するかを調べたところ、性染色体のX染色体上に多く見られ、かつ比較的広い領域(最大9Mb=900万塩基長)に固まって存在していることがわかったのである(画像4・5)。
X染色体上では、同様の特徴を持つ領域が全部で16箇所見つかり、これらは「Large Hypomethylated Domain(LoD)」と名付けられた。一般に、マウスなどほ乳類のDNAはその大部分が高メチル化されており、その中に数Kb(数1000塩基長)の小さな低メチル化領域「CpGアイランド」が点在することが知られていた。しかし、LoDのような広い領域にわたる低メチル化領域の発見は、今回が初めてである。
画像3は、マウスX染色体上のLoD領域の同定を示したものだ。体細胞と生殖細胞ゲノムのメチル化レベルが比較され、生殖細胞で低メチル化状態(体細胞の2分の1以下)にあるゲノム領域(LoD)が水色で示され、そこに含まれる遺伝子名が表示されている。がん精巣抗原遺伝子群(「Ssx」、「Fthl」、「Mageb」、「Magea」)を含む領域を*で示した。LoDはマウスX染色体上には16箇所存在し、その平均サイズは約120万塩基対。横軸はマウスX染色体を塩基対長で表してある(Mbp=100万塩基対長)。
画像4は、LoD領域のDNAメチル化、ヒストン修飾「H3K9ジメチル化」、遺伝子発現パターンの模式図。LoD領域(↔)には、生殖細胞で高発現を示す遺伝子群が集中しており、生殖細胞では低DNAメチル化状態、体細胞では高メチル化状態にある。ヒストンH3K9ジメチル化の修飾は、通常遺伝子発現に抑制的に働くといわれているが、生殖細胞、体細胞ともLoD領域に連続しており、LoDが独特なエピゲノム状態にあることがわかる。

|

|
|
画像3(左):マウスX染色体上のLoD領域の同定。画像4:LoD領域のDNAメチル化、ヒストン修飾(H3K9ジメチル化)、遺伝子発現パターンの模式図 |
|
研究チームが「興味深いこと」とするのが、LoDには生殖細胞特有に発現する遺伝子が集中して存在し、その中には「がん精巣抗原」遺伝子も複数含まれている点だ。通常、DNAの高メチル化は遺伝子発現を抑制するので、LoD領域に見られるDNAの低メチル化がこの領域の遺伝子発現を促進するのに必要であることが示されたのである。さらに、LoDの特徴として、DNAの低メチル化以外にヒストン修飾も特殊で、ほかの領域では見られない独特なエピゲノム状態にあることがわかった。この独特なエピゲノム状態が生殖細胞やがん細胞に特有な遺伝子発現の基盤になっていることが示唆された。
次に研究チームは、LoD領域がマウスから作製した幹細胞株に存在するかどうかについて探索を実施。すると、ES細胞、生殖細胞の元となる細胞から作製したEG細胞にはLoDは存在しなかった。しかし、京都大学大学院 医学研究科の篠原隆司教授らが、新生雄マウスの精巣から作製された精原細胞(精子幹細胞)に由来する幹細胞株である「GS細胞(生殖系列幹細胞)」は、雄マウスの生殖細胞と類似した遺伝子発現パターンを示し、LoDを含め、ゲノム全体で非常によく似たDNAメチル化パターンも示した(画像6)。従って、GS細胞は、生殖細胞特有な遺伝子発現とエピゲノムの関連を解析するための非常に有用な細胞リソースであることが明らかとなった。
画像5は、生殖細胞、幹細胞株、体細胞組織のDNAメチル化パターンと細胞タイプの分類。各種細胞サンプルのDNAメチル化パターンを解析し、その類似性を基に分類したものだ。胎児内の生殖細胞は、17.5日雄生殖細胞を除き、類似したメチル化パターンを持つ。多能性幹細胞であるES細胞、EG細胞も類似したメチル化パターンを示す。GS細胞のDNAメチル化パターンは新生雄マウスの生殖細胞(精原細胞)のパターンと非常に近いことがわかる。

|
|
画像5。生殖細胞、幹細胞株、体細胞組織のDNAメチル化パターンと細胞タイプの分類 |
今回、100個程度の細胞からでも解析可能な技術を開発し、生殖細胞に特有なDNA領域のLoDが発見された。今後LoDは、生殖細胞とがん細胞に共通のエピゲノムの形成や、がん細胞において生殖細胞特有の遺伝子が発現する理由などの意義について調べる手がかりになるだけでなく、生殖細胞やがん細胞のエピゲノムマーカーとしても利用できるという。
また、今回開発された解析技術はこれまで解析困難であった現象にも適用可能だ。例えば、近年、ES細胞やiPS細胞などの多能性幹細胞の集団中には、分化状態の異なる細胞集団が混在することが示されているが、こうした不均一性をもたらすエピゲノム状態の解析などへの展開が期待できるとしている。