



今回紹介する研究の主人公は、学校の教科書で紹介されることもあるテトラヒメナである。ゾウリムシと同じくくりで聞き覚えがある人もいるのではなかろうか。
このテトラヒメナやゾウリムシといった、100μm程度の繊毛虫と呼ばれる遊泳微生物は淡水中においてとても繁栄している生物群であり、虫眼鏡と少しの経験があれば、スズメやハトよりも見つけやすいという。
-
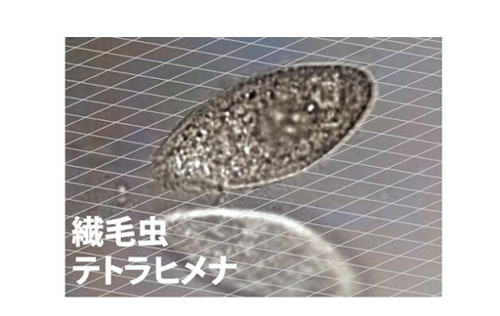
テトラヒメナの顕微鏡写真(ガラス面を識別できるように画像加工済み)(出典:京都大学)

そんな容易に見つけることができ、極小なうえに私達の生活に何ら影響を及ぼさないように思える単細胞生物でも、侮ってはならない。どれだけ小さく滑稽なものに思えていても、実は大きな世界と繋がっている、あるいは大きな循環において重要な役割を担っている可能性もある。
京都大学理学研究科、バーゼル大学、北海道大学電子科学研究所、東北大学大学院医工学研究科、基礎生物学研究所らの研究グループは、単細胞の繊毛虫テトラヒメナが水中の構造物付近で走流性を示す機構を明らかにした。走流性とは水の流れに逆らって遡上する性質の事だ。詳細は、科学ジャーナル「Science Advances」に掲載されている。
冒頭でも触れたが、テトラヒメナやゾウリムシのような繊毛虫は、一見、私達の生活に何ら影響しないように思える。しかし、実際は環境中のバクテリアを食べたり、魚の餌となり間接的に私達の食生活や地球環境を支えているのだ。
もし、繊毛虫が川から海に流され、淡水中からいなくなると、水は濁り、魚は飢え、私達の食生活を脅かすこととなる。
ここで、重要なのが海に流されていないという点だ。体長や厳しい水流にさらされるという観点から考えるに、遊泳繊毛虫は海へと流されるのではないかと思われる。
しかし、淡水中で生息しているということは、逆説的に水の流れに逆らう走流性という性質を有しているのではないかと言われてきた。
しかしながら、どの程度の流れに、どのようにして逆らうのかは解明されていなかったため、同研究グループは、テトラヒメナに人為的な流れを加えた際の挙動観察と、流体シミュレーションによって走流性のメカニズムの解明に挑んだ。
-
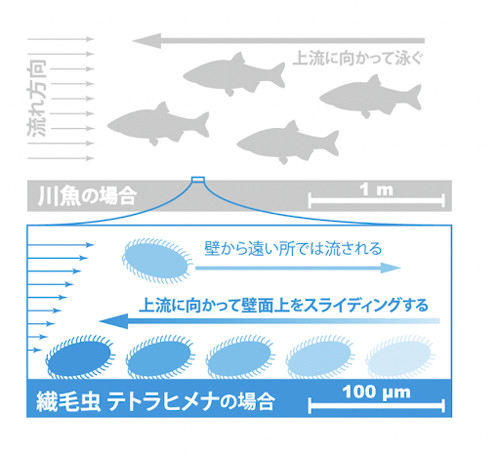
同研究の模式図(出典:京都大学)

流速の空間変化の大きさを制御できるマイクロ流体デバイスを用いて、流れ場中のテトラヒメナの挙動を定量的に評価した結果、壁付近でテトラヒメナの明確な走流性が示された。さらに、この走流性は流速の空間変化の大きさに依存していることが確認された。
-
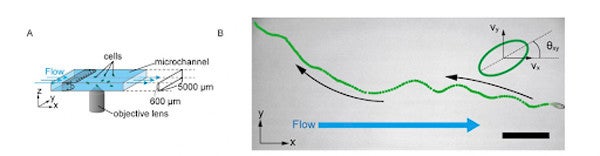
実験の模式図と流れに逆らって遡上するテトラヒメナの軌跡(論文より)(出典:京都大学)

すなわち、流れが無いときにはテトラヒメナの泳ぐ方向はランダムであるが、流れが強くなるにつれてテトラヒメナの細胞の向きが上流方向を指すようになったのだ。
次にテトラヒメナがどのように、運動しているのかを観察するために、流動場中での1本1本の繊毛運動を観察した。この観察の結果、壁付近の繊毛は運動が阻害されていることが明らかとなった。
以上の結果を、流体シミュレーションによって検証したところ、細胞形状が球状の場合では、上記の要素であっても走流性を示さない一方、テトラヒメナのような回転楕円体では、計算機内でも走流性が再現されることが分かった。
これらのことから、テトラヒメナの走流性は細胞が考えて行動しているわけではなく、細胞の形状と繊毛の性質によって決定される「カラクリ」であることが分かったのだ。
研究グループは今後、同研究成果を基礎とすることで、計算機シミュレーションなどによる生息分布予測や、それに伴う環境変化の予測の精度が改善することが期待されるとした。
それらを通じて、「気候変動」や「海の豊かさ」、「陸の豊かさ」といった持続可能な開発目標(SDGs)の実現に貢献できると考えられるのだという。















