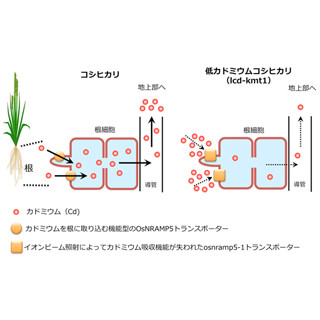
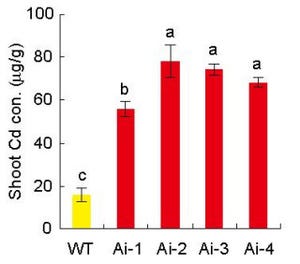
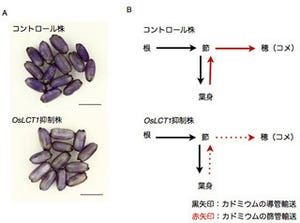
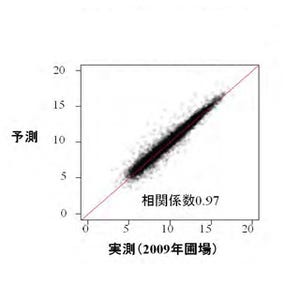
東京大学(東大)は、稲穂にできるコメ粒の数を決定する遺伝子を発見し、「TAWAWA1」と命名したことを発表した。
同成果は、日本学術振興会の吉田明希子 特別研究員PD、東京大学大学院農学生命科学研究科生産・環境生物学専攻 修士課程の笹尾真史氏(当時)、同 安野奈緒子 特任研究員、同 大門靖史 特任研究員、同 Ruihong Chen 特別研究学生、同 山崎諒 特任研究員、同 徳永浩樹 特任研究員、同大学農学部 応用生物学専修の北口善教 氏(当時)、同大大学院農学生命科学研究科生産・環境生物学専攻の経塚淳子 准教授、岡山大学 資源植物科学研究所の前川雅彦教授らによるもの。詳細は「米国科学アカデミー紀要(Proceeding of the National Academy of Science USA:PNAS)」に掲載された。
稲の穂の発生では、まず、枝梗と呼ばれる枝分かれがつくられ、その後にすべての枝に花がつき、それぞれの花が1粒のコメとなるため、枝分かれが多く形成されると1つの穂につくコメの数も増加することとなる。生物の発生過程の進行は、多数の遺伝子の協調的な作用により厳密に制御されており、こうした稲の穂の発生における、枝つくりから花つくりへのプログラム進行も同様であることが考えられているが、その制御の実態はよく分かっていなかった。
また、このプログラム進行のタイミングは稲穂につくられるコメの数に直接的に影響するはずである。これまで、花の発生に関する研究は多く行われたきたほか、近年では、作物の収量に関する遺伝子レベルでの研究も進められ、収量を左右する遺伝子の単離も報告されるようになってきた。しかし、花の発生メカニズムと収量を直接つなぐような遺伝子レベルでの知見はほとんどないのが実情であった。
そこで今回の研究では、枝分かれつくりから花つくりへと発生プログラムが転換するタイミングを決定する仕組みを解明するために、稲の変異体の探索を実施した。その結果、優性に遺伝する2つの突然変異体を発見。これらの変異体で変異を起こしている遺伝子は同一であり、どちらも優性に遺伝することが確認された。また、これら2つの変異体では、枝つくりが過剰になっており、弱い異常を示す変異体(taw1-D2)ではコメの数が増加し、異常が強い変異体(taw1-D1)では枝つくりが無限に繰り返されることを確認。この突然変異の原因となった遺伝子を「TAWAWA1(TAW1)」と命名したという。
研究グループでは、これらの変異体が稲にもともと存在するトランスポゾンにより引き起こされている可能性があると考え、トランスポゾンの転移を指標にTAW1遺伝子を単離したところ、TAW1遺伝子からは機能未知のタンパク質がつくられることが明らかになった。ちなみに稲ゲノムには、TAW1類似遺伝子がTAW1以外にも9個存在するほか、TAW1遺伝子は稲以外の植物にも広く存在する遺伝子だという。
詳細な調査の結果、2つの突然変異体(taw1-D1/taw1-D2 )ではTAW1遺伝子の制御領域にトランスポゾンが挿入されており、そのためにTAW1遺伝子の働きが高まっていることが示された。また、九州大学農学研究院の熊丸敏博 准教授らのグループとの共同研究にて、TAW1遺伝子の働きが低下した変異体(taw-3)を発見。この変異体では、枝分かれが少なく、コメ数が少ない小さな穂がつくられることが確認された。これらの結果により、穂の枝分かれの程度が、TAW1の働き程度に依存して決定されることが確認された。
さらに研究グループは、TAW1遺伝子がいつどこで働くのか(mRNAがつくられるか)を調査。その結果、TAW1 mRNAは稲の成長初期から穂の発生過程の初期まで茎頂分裂組織でつくられ、枝つくりから花つくりに転換する時点でmRNAがつくられなくなったことから、TAW1は稲の花つくりへのプログラムの進行を抑える遺伝子であると結論付けた。
なお研究グループでは、TAW1の働きがわずかに昂進したtaw1-D2変異体をコシヒカリと交配させ、taw1-D2変異を有するコシヒカリを作製。このtaw1-D2コシヒカリでは1穂のコメ数が顕著に増加し収量が増加することが確認されたほか、食味には影響がないことも併せて確認された。
今回、taw1-2変異体は人為的に作成したものではなく自然に起こった変異であり、その増加部位や程度のさじ加減が収量に対して影響をおよぼすことが示されたことから、研究グループでは、稲以外の作物にも存在するTAW1遺伝子を活用することで、将来的に種子や果実を収穫する作物の収量の増加につなげることが期待できると説明している。

|
|
左が通常のコシヒカリ、右がTAW1コシヒカリ |