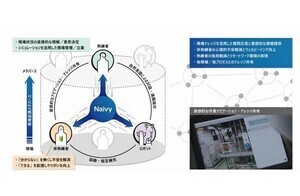
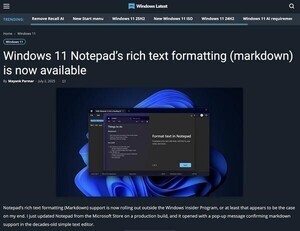
理化学研究所(理研)は、免疫応答の核となるT細胞の活性化や増殖を抑制する補助刺激受容体「PD-1(Programmed Cell Death-1)」が、「ミクロクラスター(集合体)」を形成し、T細胞の過剰な活性化を抑制していることを明らかにしたと発表した。
成果は、理研免疫・アレルギー科学総合研究センター 免疫シグナル研究グループの斉藤隆グループディレクター(副センター長、大阪大学免疫学フロンティア研究センター招へい教授兼務)、横須賀忠上級研究員(科学技術振興機構さきがけ研究員兼務)と、大阪大学免疫学フロンティア研究センター、東京医科歯科大学大学院医歯学総合研究科らの共同研究グループによるもの。研究の詳細な内容は、日本時間5月28日付けで米科学雑誌「The Journal of Experimental Medicine」に掲載された。
また、今回の研究成果は、科学技術振興機構 戦略的創造研究推進事業 個人型研究(さきがけ)「炎症の慢性化機構の解明と制御」研究領域における研究課題「MAPK経路の分子イメージングによるT細胞活性化遷延機構の解明」の一環として行われたものである。
生体を外敵から守る免疫応答は、まずウイルスや花粉などの異物が「抗原提示細胞」によって取り込まれ、抗原としてT細胞に提示されることから始まる。その後、抗原を認識したT細胞が活性化し、さまざまな情報伝達物質を放出したり、がん細胞やウイルス感染細胞を殺したり、抗体産生を促すなど、より特異的で高度な免疫応答を長期にわたり引き起こす。
抗原提示細胞とは、免疫細胞の内、外から侵入した病原体や体内で生じた死細胞などを貪食し、抗原として自らの細胞表面上に提示する能力を持つ細胞を示す。代表的なものにマクロファージや樹状細胞などがある。抗原提示細胞は、外来抗原によって自らも活性化する(自然免疫)と共に、T細胞やB細胞を活性化し、より特異的で高度な免疫応答(獲得免疫)を誘導する。
抗原認識の際、T細胞は抗原提示細胞と強く接着し、その接着面には、T細胞表面の受容体を中心に、接着分子である細胞内のシグナル伝達分子が同心円状に並ぶ。この構造は、神経細胞のシナプスに似ていることから、「免疫シナプス」と呼ばれるようになった(画像1・2)。
免疫シナプスの直径は約10μmで、中心部を「Central-supramolecular activating cluster(c-SMAC)」、接着分子からなる周辺部を「Peripheral-SMAC(p-SMAC)」と呼ぶ。B細胞やナチュラルキラー細胞など、細胞間相互作用を行う免疫細胞の接着面に広く見られるのが特徴だ。
2005年に免疫シグナル研究グループは、免疫シナプスの中に、T細胞受容体と、リン酸基を付加する「酵素キナーゼ」やタンパク同士の結合に必要なアダプターなどの「シグナル伝達分子」から構成される分子の集合体「ミクロクラスター」が存在し、これがユニットとして、T細胞活性化の開始と維持を行うことを明らかにしていた。
画像1は免疫シナプスの構造とミクロクラスターの模式図(初期のT細胞活性化とT細胞活性化の維持)で、画像2はその蛍光顕微鏡像だ。画像1の左側は、初期T細胞活性化の模式図。抗原提示細胞がT細胞に結合した際と、結合部分で形成されるミクロクラスターを拡大したものである。
T細胞が抗原提示細胞に接着すると、T細胞受容体が抗原提示細胞上の抗原と結合し、T細胞受容体を核とするシグナル伝達分子のミクロクラスターが形成される。
このミクロクラスターには、T細胞受容体下流のシグナル伝達分子が会合し、T細胞活性化シグナルを伝える活性化ドメインとしてT細胞の初期の活性化を引き起こす。
正の補助刺激受容体「CD28」は、CD28の「リガンド」と結合することでミクロクラスターに集まり、CD28の下流に特異的な別のキナーゼを介してより強力なシグナルを伝える仕組みだ。なおリガンドとは、受容体(レセプター)が結合する相手方の分子のこと。受容体はリガンドと結合することで、受容体から、細胞質や核にある次の分子へと情報が伝達される。
そして、その画像1の右側の図は、左側から5分が経過した、T細胞活性化の維持を表した模式図。初期の接着から5分が経過すると、T細胞受容体は接着面の中央部に移動、T細胞受容体下流のシグナル伝達分子は解離し、活性を失う。CD28はT細胞受容体の周囲に輪状構造を構築し、CD28下流のキナーゼを留まらせることにより、T細胞受容体シグナルとは別の活性化シグナルを伝える仕組みだ。
また、接着面の周縁部では新たなミクロクラスターが繰り返し形成され、T細胞受容体からのシグナルを伝え続ける。T細胞受容体とCD28からなる接着面の中心を「Central-supramolecular activating cluster(c-SMAC)」、接着分子からなる周辺部を「Peripheral-SMAC(p-SMAC)」といい、この同心円状構造を免疫シナプスと呼ぶ。
画像2の蛍光顕微鏡像は、画像1の2つの模式図のそれぞれの蛍光顕微鏡画像。擬似的な抗原提示細胞膜である「プレイナーメンブレン」に載せたT細胞である。T細胞受容体(赤)、接着分子(水色)、CD28下流で働くキナーゼ(緑)。

|

|
|
画像1(左)は、免疫シナプスの構造とミクロクラスターの模式図で、画像2はその蛍光顕微鏡像 |
|
T細胞の活性化には、T細胞受容体ミクロクラスターからのシグナルのほかに、補助刺激受容体からのシグナルがバランスよく伝わることが必要と考えられている。T細胞は、T細胞受容体からの刺激で活性化するが、それ単独の刺激だけでは、細胞死や不応答を起こしてしまうのを防ぐためだ。
そこでT細胞に発現するのが、T細胞受容体以外のさまざまな補助刺激受容体である。補助刺激受容体は、T細胞受容体からのシグナルと協力し、T細胞の活性化、分化、増殖を調節する役目を担う。補助刺激受容体には、T細胞の活性化に対し正の働きをするものと、負の働きをするものとが知られている。
免疫シグナル研究グループはこれまで、正の補助刺激受容体であるCD28もミクロクラスターとなり、特殊なキナーゼと共に免疫シナプス中心部で新たな活性中心を作り、T細胞の活性化を増強していること、また負の補助刺激受容体「CTLA-4」もミクロクラスターとなり、CD28の活性中心を破壊することでT細胞の活性化を抑制していることを明らかにしてきた。
近年、もう1つの負の補助刺激受容体PD-1が、より広範にT細胞の活性化を抑制する分子として注目されている。PD-1はI型膜タンパク、免疫グロブリンスーパーファミリー分子の1つで、細胞死誘導時に発現が増強されるタンパク質として、1992年に京都大学本庶佑研究室にて単離同定された。活性化したT細胞、B細胞、マクロファージに発現しており、リガンドには「PD-1ligand1(PD-L1)」と「PD-L2」の2つが知られている。
PD-1の遺伝子を欠損したマウスが、ループス腎炎や関節炎、拡張型心筋症などの全身性自己免疫疾患により死亡することから、PD-1は長期的なT細胞の活性化抑制に重要であるという推測がなされた。
慢性C型肝炎やエイズウイルス感染などの慢性炎症の際に出現する「消耗T細胞」もPD-1を高発現しており、PD-1のリガンドへの結合を阻止することで消耗T細胞の機能が回復することから、PD-1の中和抗体も臨床応用されつつある。
なお消耗T細胞とは、長期的な感染や担がん状態のように抗原刺激が慢性的に及んだ場合、本来反応すべき外来抗原に対して無反応になってしまったT細胞のことをいう。消耗T細胞はPD-1を高発現しており、PD-1のリガンドへの結合を阻害すると、T細胞の反応性が回復することが知られている。
またPD-1は、加齢に伴うT細胞免疫応答性の低下を示す「免疫老化」のマーカー分子であることも示された。しかし、臨床的な有用性が先行しているのに対し、PD-1を介するT細胞抑制メカニズムの解明は進んでいない。
これまでは、マウスによる個体レベルでの解析や細胞を用いた生化学的解析が中心で、分子レベルの解析は行われていなかった。そこで研究グループは、PD-1がT細胞の活性化をどのように抑制するか、分子レベルの解析に挑んだというわけだ。
T細胞でのPD-1分子のリアルタイムな動きを、「分子イメージング」技術を用いて解析するため、まずT細胞受容体のリガンドである主要組織適合抗原複合体とPD-1のリガンドであるPD-L1をガラス平面上の人工脂質二重膜に載せた、擬似的な抗原提示細胞膜「プレイナーメンブレン」が開発された(画像2)。
次にプレイナーメンブレン上にT細胞を置き、T細胞とプレイナーメンブレンとの接着面で起きる現象を、緑色蛍光タンパク質などを付加させた分子を指標にし、全反射蛍光顕微鏡や共焦点レーザー顕微鏡で観察した。
ちなみに分子イメージングとは、細胞内や生体内の分子を可視化する技術のことをいう。蛍光を用いて、蛍光顕微鏡やレーザー顕微鏡で観察する場合が多い。着目している分子と蛍光タンパク質との融合タンパク質を細胞内で発現させたり、蛍光物質で標識した抗体を用いたりするため、細胞内外の分子の動きを、細胞が生きたまま観察できる点が大きな特徴だ。
蛍光タンパク質や蛍光物質には、発光波長の違いによりさまざまな色がある。緑色蛍光タンパク質であるGFP(Green Fluorescence Protein)が基本になっているが、複数の色を組み合わせることにより、多数の分子を同時に観察することも可能だ。
画像3は、プレイナーメンブレン法によるT細胞受容体及び細胞内分子の観察方法の模式図。実際の細胞は球形なのはいうまでもないが、ここでは円として描かれており、また細胞膜の複雑な凸凹も省略されている。
プレイナーメンブレン法では、抗原提示細胞に発現している主要組織適合抗原複合体や接着分子などの細胞表面分子に膜結合アンカー配列を付加し、カバーガラス上に作製した平らな人工脂質二重膜に導入。そして、この抗原提示細胞の細胞膜を疑似したプレイナーメンブレン上にT細胞を落下させ、蛍光顕微鏡で細胞内外の分子の挙動を観察するという流れだ。
観察したい分子は、蛍光で標識された抗体で染色したり、直接蛍光物質を付加したりする。全反射蛍光顕微鏡はレーザーの入射角を大きくして微妙に漏れ混む「エバネッセンス(エバネッセント)光(近接場光)」を利用するため、カバーガラスから100ナノメートル(nm)といった細胞膜近傍の分子の観察が可能だ。
また、このような弱い光を用いてバックグラウンドを下げることで、1分子レベルでの解析も可能にしている。さらに共焦点レーザー顕微鏡では、細胞表面から細胞内部までの分子の挙動を、幅広い波長の蛍光を用いて観察することが可能だ。

|
|
画像3。プレイナーメンブレン法によるT細胞受容体及び細胞内分子の観察方法の模式図 |
観察の結果、T細胞がプレイナーメンブレンに接着すると、ただちにT細胞受容体はミクロクラスターとして集まり、集まったT細胞受容体ミクロクラスターは数分の内に免疫シナプスの中心に移動、凝集することが判明した。
同時に、PD-1もミクロクラスターを形成し、免疫シナプスの中心に移動し、凝集したのである(画像4)。また、PD-1はミクロクラスターを形成する際、数10秒という短時間で、脱リン酸化酵素「SHP2」をミクロクラスターに呼び込み会合すること(画像5)、そしてこのSHP2がミクロクラスターに集まるシグナル伝達分子を直接脱リン酸化して、T細胞受容体からの活性化シグナルを抑制することもわかった。
なお脱リン酸化酵素とは、リン酸化されたチロシン残基、スレオニン残基、セリン残基などからリン酸基を外す酵素のことだ。リン酸基を取り除くことは、シグナルの遮断を意味する。
画像4は、T細胞受容体とPD-1ミクロクラスターの蛍光顕微鏡観察像。上のモノクロ画像は、蛍光タンパク質を付加したPD-1をT細胞に遺伝子導入後、プレイナーメンブレン上に落下させ、全反射蛍光顕微鏡を用いてリアルタイム観察を行ったもの。PD-1はリガンドがあるとクラスターを形成し、数分以内に中央部(c-SMAC)に集まった。
下の画像は、共焦点レーザー顕微鏡を用いた、T細胞のT細胞受容体(赤)とPD-1(緑)のリアルタイム観察結果。T細胞とプレイナーメンブレンとの接着1分後、T細胞受容体とPD-1のミクロクラスターは同じ場所に形成される(黄)。20分後、T細胞受容体もPD-1も接着面の中央に集まり、c-SMACを形成した。

|
|
画像4。T細胞受容体とPD-1ミクロクラスターの蛍光顕微鏡観察像 |
そして画像5は、PD-1ミクロクラスターに集まるSHP2の蛍光顕微鏡画像。上の列は、蛍光タンパク質を付加したPD-1(黄)とSHP2(水色)をT細胞に導入後、プレイナーメンブレン上に落下させ、共焦点レーザー顕微鏡を用いてリアルタイム観察を行ったもの。 PD-1とSHP2のミクロクラスターが同じ部位に存在するのが観察され、「蛍光共鳴エネルギー移動(FRET :Fluorescence resonance energy transfer)」においても、SHP2とPD-1の会合が示された。
蛍光共鳴エネルギー移動とは、近接した2個の色素分子の間で、励起エネルギーが電子の共鳴により直接移動する現象のこと。一方の分子(供与体)に吸収された光のエネルギーによって、他方の分子(受容体)にエネルギーが移動し、受容体から蛍光が放射される。2つの分子の数nm以内の接近を意味する。
画像5の下の列は、同T細胞を抗原提示細胞と接着させ、2つの細胞の間に集まるPD-1(黄)とSHP2(水色)を、共焦点レーザー顕微鏡を用いて観察したものだ。

|
|
画像5。PD-1ミクロクラスターに集まるSHP2 |
さらに、活性化シグナルを失ったT細胞は、免疫シナプスを保持できず、プレイナーメンブレン上を無作為に動き回り始め、その結果、T細胞の活性化が中断されることがわかった。
慢性炎症が続くと体内に現れる消耗T細胞がPD-1を高発現していることは前述した通りだが、消耗T細胞がPD-1のミクロクラスターを作ること、PD-1のリガンドへの結合を阻害すると、PD-1のミクロクラスターは消失し、消耗T細胞の機能が回復することも確認された。
これまで、PD-1もCTLA-4も、T細胞の活性化を抑制する負の補助刺激受容体であることは知られていたが、先行研究と今回の成果により初めて両者のT細胞活性化抑制メカニズムの違いを明らかにすることができたのである(画像6)。
画像6は、PD-1とCTLA-4ミクロクラスターによるT細胞活性化抑制を表した模式図だ。左側の「初期のT細胞活性化」は、正常な初期のT細胞活性化が、T細胞と抗原提示細胞との接着面に形成されるT細胞受容体-CD28のミクロクラスターによって引き起こされ、主にキナーゼやアダプターなどのシグナル伝達分子が働いていることを表している。
PD-1はリガンドと結合することでT細胞受容体とミクロラスターを形成し、そこに集まるキナーゼによりPD-1自身がリン酸化を受け、SHP2をミクロクラスターに呼び寄せる仕組みだ。次にSHP2は、T細胞受容体-CD28ミクロクラスターに集まるシグナル伝達分子を脱リン酸化し、ミクロクラスターからの活性化シグナルを減弱、つまり最初の活性化反応に対する負の制御を行う。
右の模式図は、「T細胞活性化の維持」を表したもの。T細胞活性化の維持は、主にc-SMACに形成されるCD28とその下流のキナーゼからなる活性化中心によって行われる。CTLA-4はCD28からリガンドを奪い取ることで、CD28の活性中心を破壊し、CD28シグナルによるT細胞活性化を抑制する仕組みだ。
PD-1、CTLA-4の2つの負の補助刺激受容体は、時間的、空間的に別の方法で、異なる活性化シグナルを抑え、二重にT細胞の活性化を制御するのである。

|
|
画像6。PD-1とCTLA-4ミクロクラスターによるT細胞活性化抑制 |
以上から、T細胞の活性化だけでなく、T細胞を抑制するミクロクラスターがあることが判明した。今回の研究は、T細胞活性化を調節する分子のダイナミズムを観察することで、免疫応答の亢進と抑制の新たな分子メカニズムを解明したものといえよう。
また、PD-1ミクロクラスターの存在を明らかにし、その抑制のメカニズムも今回の研究によって解明された。CTLA-4を初めとする負の補助刺激受容体はいくつか存在するが、それぞれが異なった調節機構を持ち、多方面からT細胞の過剰な活性化を効率よく制御していることが示唆される。
現在、慢性C型肝炎やエイズウイルス感染を排除する、また、がん患者での免疫系を活性化するために、抗PD-1抗体や抗PD-1リガンド抗体が臨床応用され始めており、すでに臨床応用されている抗CTLA-4抗体よりも効果が顕著であるという報告がなされている状況だ。
しかし、免疫系の全身的な活性化による二次的な自己免疫疾患の発症など、阻害抗体使用による副作用も危惧されるという。
今回の成果は、PD-1の動態を制御するなど新しい観点からの創薬と共に、より選択的な免疫抑制剤や免疫活性剤の開発への可能性を示しており、安全で効果的な免疫治療の進歩につながると期待できると、研究グループはコメントしている。