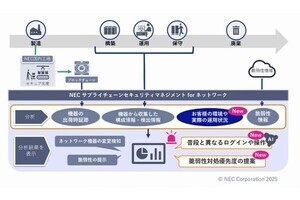
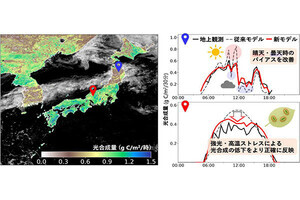
理化学研究所(理研)は1月16日、ショウジョウバエにおいて、気管上皮細胞内の「逆行性小胞輸送システム」が気管の長さを制御していることを明らかにし、管の長さと太さという2つの形状特性が、異なるタンパク質輸送システムによって制御されていることが示されたと発表した。
成果は、理研 発生・再生科学総合研究センター 形態形成シグナル研究グループの董波 国際特別研究員、同・林茂生グループディレクターらによるもの。研究の詳細な内容は、日本時間1月16日付けで英国のオンライン科学雑誌「Nature Communications」に掲載された。
進化の過程において陸生生物が大型化していくには、体内で血液、空気、水分などを効率的に循環させることが必要だ。それを可能にするために、動物の血管や呼吸管、植物の師管や導管などの管状組織のネットワークが縦横に張り巡らされ、高度に発達してきたと考えられている。
効率のよい循環を維持するには、それぞれの個体や組織のサイズ、さらに管内を流れる物質の物理的性質などに合わせて、管の長さや太さを適切に調整することが必要だ。
脊椎動物の場合、血管の太さや長さは心臓が動き始め血流が流れる前におおむね決まっているため、発生過程のある段階で管の形状を決める仕組みが細胞や組織に備わっていると考えられるという。
過去の研究からショウジョウバエの気管の太さは、気管の内側(管腔)を占めるキチン質を主成分とした「細胞外基質」が蓄積することで制御され、長さについては「キチン修飾酵素」によってキチン質が化学修飾を受けることで、過剰に伸長しないように制御されていることが知られている。なおキチン修飾酵素は、気管を形成している「上皮細胞」内で合成され、「小胞輸送システム」を経由して細胞外へ分泌される仕組みだ。
小胞輸送システムには、輸送される分子に応じて多様な経路が存在することが知られている。通常、細胞内で合成された分泌タンパク質は、いったん小胞体内へ運ばれ品質管理を受け、正常に機能するものだけが「ゴルジ体」へと運ばれる形だ。
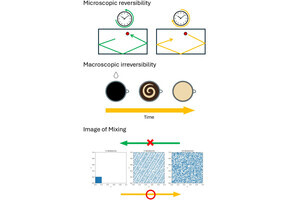
ゴルジ体へ運ばれたタンパク質は、糖鎖の修飾や輸送先への選別が行われ、選別されたタンパク質は輸送小胞に詰め込まれ、目標の細胞内小器官、細胞膜や外へ運ばれる。この順方向の輸送システムに対して、細胞外から取り込まれたタンパク質がゴルジ体に戻される経路があり、「逆行性小胞輸送システム」という。このようなシステムを通じて行われる細胞と外部環境との分子のやり取りは、生物共通の生命維持活動に必須のシステムだ(画像1)。
細胞外から取り込まれたタンパク質は小胞体(「初期エンドソーム」、「後期エンドソーム」)で選別を受け、ただちに細胞外に放出されたり、逆行性小胞輸送経路を通じてゴルジ体に運ばれ、再修飾した後に細胞外に分泌される。また不要なタンパク質が「リソソーム」で分解される経路もある。

|
|
画像1。小胞輸送システム |
ショウジョウバエ気管の太さ制御には、「アクチン重合因子」を介したタンパク質輸送システムが関わることが知られていた。しかし、長さ制御の仕組みについては、管腔に分泌されるキチン修飾酵素が必要だとわかっていたものの、タンパク質輸送システムがどのようにキチン修飾酵素の量の調節に関与しているのか、その細胞内の仕組みについては不明のままだったのである。
そこで研究グループは今回、管状組織を形作るための細胞内の仕組みを解明するために、ショウジョウバエの変異体を用いて、キチン修飾酵素の調節に関わる要因の探索に挑んだ。
具体的には、逆行性小胞輸送システムに関わる「rab9遺伝子」の変異体を調べたところ、気管が過剰に伸長する表現型を示すことが見出された(画像2)。
画像2は、rab9遺伝子変異体は過剰な気管伸長を示す証拠の画像。ショウジョウバエ胚の気管の管腔に存在するキチン質が含まれた細胞外基質を染色している。rab9遺伝子変異体は前後に走る太い管が過剰な気管長を示す(下図)。太さは正常胚(上図)と変わらない。

|
|
画像2。rab9遺伝子変異体は過剰な気管伸長を示す |
気管の太さと配置は標準と変わらず、胚発生自体はおおむね正常に進行していたので、気管形成の内、長さに関わる機能だけが阻害されていると考えられた。そこで、rab9遺伝子変異体のキチン修飾酵素の分布を経時的に調査したところ、キチン修飾酵素は、中期胚(受精後10時間目)における気管発生時には管腔内に分泌されるにも関わらず、後期胚(受精後15時間目)になると気管の管腔内から著しく減少していることが確認された。これは、気管の管腔に存在する分子の内キチン質やほかのタンパク質の分布は正常だったことから、キチン修飾酵素を輸送するシステムだけに異常が生じていることが考えられたという。
画像3は、rab9遺伝子変異体で見られるキチン質修飾酵素の分布異常を示したもの。rab9遺伝子変異体の後期胚(受精後15時間)では、キチン修飾酵素(Serpはキチン質修飾酵素をコードしている遺伝子のこと)の分布が管腔から減少する。一方で太さ制御に関わるタンパク質(2A12、Pio)やキチン質(Chitin)の分布に変化はなかった。

|
|
画像3。rab9遺伝子変異体で見られるキチン質修飾酵素の分布異常 |
さらに、Rab9タンパク質に結合するほかの分子を生化学実験で調べたところ、逆行性小胞輸送システムに関わるタンパク質「Vps35」がRab9タンパク質と結合することが明らかになったのである。
そこで、正常なショウジョウバエの培養細胞を用いて、Rab9タンパク質の動きをライブイメージングで詳細に観察を行った。その結果、これらは小胞体内で局所的な集合体を作り、小胞体膜を細胞質側に突出させ、さらにアクチン重合に関わるタンパク質「WASH」とアクチン繊維の働きで小胞体の一部が切り取られる様子が観察された。
画像4は、Rab9小胞の変形と切り出し。Rab9タンパク質で標識された小胞(ピンク)は、正常なショウジョウバエの培養細胞中で突起構造を形成し、アクチン繊維(緑)と共局在し、細胞質側へ(白矢じり)切り出される。

|
|
画像4。Rab9小胞の変形と切り出し |
重要なことに、この切り出された小胞体断片にはキチン質修飾酵素が含まれていた。そのため、一度分泌されたキチン質修飾酵素が細胞外から取り込まれ、小胞体内で選り分けられ、Vps35とRab9タンパク質の働きで小胞体断片として切り出され、ゴルジ体へ運ばれ再修飾を受け、活性化型として細胞外分泌の経路に送られると推察されたのである。キチン修飾酵素は、このように再利用され管腔での酵素の働きを維持することで、気管が異常に伸長しないように制御していることが示唆された。
画像5は、小胞体の切り出しによるキチン修飾酵素の選別。細胞内に取り込まれたキチン修飾酵素「Serp」(オレンジ色の粒)は小胞体に移行し、Rab9タンパク質と逆行性小胞輸送システムに関わるタンパク質Vps35によって形成される細部膜突起に集められる。この細胞膜突起構造はアクチン重合分子WASHの働きで重合するアクチン繊維構造によって切り出され、その後にゴルジ体に輸送されるものと考えられるという。

|
|
画像5。小胞体の切り出しによるキチン修飾酵素の選別 |
また、この逆行性小胞輸送システムを変異させた場合、長さが過剰になる一方で太さ制御に関わる多くの分子の分布は正常であったため、太さ制御にはこのシステムは関与しないことがわかった。
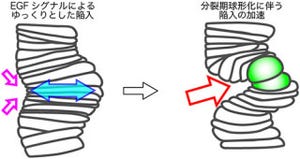
このことから、管の長さと太さの制御には別々の小胞輸送システムが関わることが明らかになったのである。つまり、異なるシステムが別々に対応していることがわかったというわけだ(画像6)。
画像6は、気管形成における2種類のタンパク質輸送システム。左側は、長さ制御に関わるタンパク質輸送システム。逆行性小胞輸送システムでは管腔から取り込まれたキチン修飾酵素Serpが小胞からゴルジ体へ送られ、再修飾を受けた後に再分泌される。Serpはキチン質を修飾してキトサン(Chitosan)に変換する。一方の右側は、太さ制御に関わるタンパク質輸送システム。「アクチン重合因子Dia依存的経路」では太さ制御に関わるタンパク質(「2A12」「Pio」)が管腔に分泌される。

|
|
画像6。気管形成における2種類のタンパク質輸送システム |
この逆行性タンパク質輸送システムは、ショウジョウバエからヒトを含む高等動物に保存されており、輸送されるタンパク質について数多く知られているが、管状組織の形を決める形態形成に関わることは知られていなかった。
今回の発見は、形態形成を起こす1つの仕組みとして非常に興味深い成果であると、研究グループはコメント。今後、さらに研究を発展させることで、ヒトを含めた脊椎動物での管形成におけるタンパク質輸送システムの役割を解明する手がかりになると期待できるとしている。