京都大学は3月4日、東京大学との共同研究により、がん化学療法の障害となっている「ATP結合カセット(ABC:ATP Binding Cassette)多剤排出トランスポーター」と非常によく似た構造と機能を示す膜タンパク質「CmABCB1」を温泉に棲む真核生物から発見し、その分子構造と多剤排出メカニズムを解明したと発表した。
成果は、京大 薬学研究科の加藤博章 教授(理化学研究所(理研)客員研究員兼任)、同・大学 物質-細胞統合システム拠点(iCeMS)の植田和光 教授、同・大学 化学研究所の平竹潤 教授、東大 理学系研究科の菅裕明 教授らの共同研究チームによるもの。研究の詳細な内容は、米国東部時間3月3日付けで米科学雑誌「米科学アカデミー紀要(PNAS)」先行オンライン版に掲載された。
生物の細胞は「トランスポーター(膜輸送体)」という膜タンパク質を用いて、細胞膜の内外の物質輸送を行っている。中でも、地上の生物のエネルギー源である「ATP(アデノシン三リン酸)」によって駆動される駆動される部位を共通に持っている一群のタンパク質分子が、ABCトランスポーターだ。
同タンパク質の仲間は、ヒトでは48種類が知られており、そのほとんどが病気と関連している。中でも、ABCBファミリー(ABCファミリーのサブグループB)に属する「P糖タンパク質」または「MDR1」とも呼ばれる、「ABCB1」は最も有名だ。ちなみにABCB1は、1986年当時、米・NIH(米国国立衛生研究所)へ留学していた今回の研究チームの一員である植田教授によって世界で最初に単離された多剤排出トランスポーターである(ABCトランスポーターファミリーに属していることは後ほど判明した)。
ABCB1は生体を外界の異物から保護する重要な分子だが、その一方でがん細胞において沢山作られており、がんの多剤耐性の原因として抗がん剤治療の障害にもなってしまっているという厄介な性質を持つ。なお、多剤排出トランスポーターの高発現もがん細胞の多剤耐性の原因となることが多い。がん細胞では、ABCB1のほか、「ABCC1(MRP1)」、「ABCG2(BCRP)」の3種類のABCトランスポーターが多剤耐性と関わると考えられている。
ABCB1は、多種多様な化学構造の化合物を細胞外へと排出する能力を持つ。ABCB1は、小腸、血液脳関門、肝臓、腎臓、生殖器に多く存在しており、外界の異物から生体を守る役割を果たしている。一方で、経口投与された薬もABCB1によって体外へと排出されることから、ABCB1との相性が薬の効力と大きく関係しているというわけだ。
がんがABCB1によって多剤耐性を獲得する仕組みは、がんを完全に死滅させられないことで起きてくる。抗がん剤は本来はよく効き、ほとんどのがんが死滅するのだが、生き残ってしまったがん細胞があると、これが多剤耐性獲得のため、ABCB1を大量に作る。すると、初回に用いた抗がん剤どころか、まだ使っていないそのほかのさまざまな抗がん剤にまで効かない多剤耐性を獲得してしまうのだ。
このことから、ABCB1の分子構造を解明してこの多剤排出という機能を解明することは、基礎科学的に重要なことはもちろん、創薬や医療にとっても重要な意義を持っているのである。このABCB1遺伝子を植田教授が単離に成功して以来、多剤排出のメカニズムを明らかにするため、世界中の研究者が結晶構造解析を30年近く試みてきた。しかし、ABCB1は細胞から取り出すと安定性が低く、詳細な分子構造の決定はこれまで不可能だったのである。
そこで研究チームは今回、ヒトのABCB1が不安定で結晶にならないのであれば、高温に棲む生物から性質のよく似た分子を探すという方法を考案。そして、「シゾン(Cyanidioschyzon merolae)」という温泉に棲む真核生物に着目し、ヒトABCB1と遺伝子配列のよく似たCmABCB1(「シゾンのABCB1」という意味で命名された)が発見されたのである。その後、京大ではCmABCB1の結晶化に成功して、その分解能を向上させ、結晶構造を決定することに成功。
また、CmABCB1がヒトABCB1とよく似た化合物排出作用を示すことについても確認された。そのほか、東大 理学系研究科によるCmABCB1の排出扉に外側から閂を掛けるような仕組みの阻害剤「aCAP」(結晶の分解能向上をもたらせもした)の開発などもされた。なお、不安定な膜タンパク質の結晶であることから、X線結晶構造解析では理研が所有し高輝度光科学研究センターが運用する播磨にある大型放射光施設「SPring-8」が利用されている。
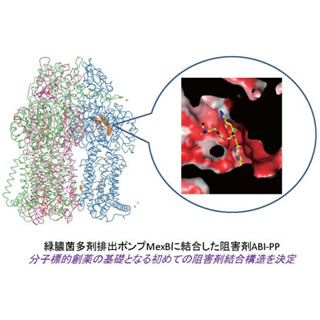
CmABCB1は、細胞膜に埋め込まれた排出ポンプ領域「膜貫通領域(TMD:trans-membrane domain)」と細胞質に浮かぶATP駆動エンジン領域「ヌクレオチド結合領域(NBD:nucleotide binding domain)」からできている(画像1)。ATPはDNAやRNAを構成するヌクレオチドの1種なので、NBDを扱えるのである。
NBDは2つの塊でできており、その中央部にATPを結合して分解する装置を持つ。そして、ATPとの結合、ATPを「ADP(アデノシン二リン酸)」とリン酸に分解、生成したADPからの解離、を1サイクルとして2つの塊が結合と解離を繰り返し動力を発生する。これが、TMDに伝わりポンプが駆動されるという仕組みだ。
画像が、1CmABCB1の結晶構造の模式図である。CmABCB1は2つのサブユニットが1組で作用する仕組みを持つ。この模式図では、一方が彩色され、他方は灰色で表現されている。彩色したサブユニットでは、TMDを構成する6本の円柱状構造(αヘリックスTM1~TM6)に番号1~6が振られており、αヘリックスはリボンで表現された。TM2(ブルーグリーンと灰色)の細胞外と細胞膜の境界付近に結合しているのは、阻害剤aCAP(黄色と黒)だ。また、細胞膜との関係がわかるように、膜を構成するリン脂質がモデルで示されている。

|
|
画像1。CmABCB1の結晶構造の模式図 |
TMDは、画像1の説明でも触れたが、6本の円柱状構造をしたαヘリックスの2組、合計12本が巧みに編み込まれることで中央に巨大な空洞を作り、その細胞外側を1番と6番のαヘリックス(TM1とTM6)2組4本で、細胞質側を3番と4番のαヘリックス(TM3とTM4)2組4本で、それぞれ開口部を作り、それらを結ぶ2番と5番のαヘリックス(TM2とTM5)2組4本で築かれている仕組みだ。
その結果、TMDには、細胞膜と細胞質が接する境界付近に横へ大きく口を開けた化合物取り込み部位ができあがり、今回の研究では、そのゲートが化合物の大きさに応じて変化できる柔らかさを持っていることが判明した。そして、その取り込み口の奥、すなわちTMDの中央部に巨大な空洞が作られ、その天井は疎水性の化合物が吸着しやすい構造であることも確認されたのである。
さらに、天井の横には天井の中央の開口部(TM1とTM6の2組)を開く引き金となるスイッチがあることも判明。TMDとNBDは短い連結部位によって結ばれており、ATPの結合、分解、解離の周期で構造変化するNBDエンジンの動作が2番と5番のαヘリックス(TM2とTM5)2組4本を介してTMDに伝わる仕組みが見えてきたという。
なお疎水性と親水性とは、化合物の水への溶けにくさ、溶けやすさのことで、水には溶け難いのが疎水性、水に溶けやすいのが親水性。また、その両方の性質を持つ「両親媒性」という洗剤のような物質もある。疎水性の化合物は、リン脂質でできている細胞膜になじみやすく細胞膜を透過しやすい。しかし、高塩濃度水溶液である細胞質には溶け難いというわけだ。
前述した構造と機能解析から導出された多剤排出のメカニズムは、次のようになるという(画像2)。細胞膜を通過しやすい疎水性の化合物は細胞質には溶け難いため、外界から細胞へと侵入して来ると一時的に細胞膜と細胞質の境界に溜まる。それをCmABCB1は大きさ自在な取り込み口で内部の空洞へと吸い込むというわけだ。
その空洞の天井は疎水性の化合物を吸着しやすいため、化学構造の違いをあまり問わず、何でも吸い込むことができると考えられるという。そして、天井近くの化合物がスイッチに触れることを引き金に天井の扉(TM1とTM6)が開き、化合物は細胞の外へ排出されるというものだ。天井および細胞質側におけるそれぞれの扉を開閉させる際の動力は、ATPエンジン(NBD)の動きが供給している。

|
|
画像2。多剤排出のメカニズム |
今回の成果により、多剤排出の基本的な仕組み、また、ABCB1に特異的に結合する阻害剤の新たな設計指針が明らかになった。これらの情報は、薬の設計合成あるいは薬物治療の改良に役立つものと期待されるという。今後、研究チームでは、CmABCB1の作用に伴う動きの全貌を解明し、ABC多剤排出トランスポーターの仕組みと弱点を詳しく解明すると同時に、CmABCB1で判明したことがヒトABCB1についても当てはまるかどうかの研究を行う予定としている。