東京大学と科学技術振興機構(JST)は7月12日、霊長類大脳皮質の階層的な領野構造で作り上げる外界の情報データベース「外界の内部表現」(内部表象)の新しい計算原理を発見したと共同で発表した。
成果は、東大大学院 医学系研究科 機能生物学専攻 統合生理学分野の宮下保司教授、同・平林敏行特任講師らの研究チームによるもの。研究は科学技術振興機構(JST)戦略的創造研究推進事業(CREST)研究領域「脳神経回路の形成・動作原理の解明と制御技術の創出」の一環として行われ、詳細な内容は7月12日付けで米科学誌「Science」に掲載された。
ヒトは外の世界を認識する際に、自分の頭の中に外の世界の写真のようなコピーを作っているわけではない。眼の前のコーヒーカップは当たり前だが、見る角度によって異なる形・大きさに見える。しかし、同じコーヒーカップであることはすぐにわかることもいうまでもない。
この仕組みは、ヒトの脳が「外界の内部表現(神経表象または内部表象)」と呼ばれる情報データベースを作り上げており、眼からの入力情報に基づいてこの情報データベースを自在に使うことによって可能になるもの(画像1)。この「外界の内部表現」は脳の領野ごとに階層的に構成され、高次の領野ほど、より複雑な特徴を表象するようになっているのである。

|
|
画像1。大脳における外界の分散型内部表現。外界の情報(ここでは、見えている馬)の具体的内容(例えば、色や形)は、大脳皮質において分散して階層的に処理・貯蔵される |
物体の視覚特徴の表象様式は大脳皮質の内部表現の中でも最もよく調べられているが、個々のニューロンの活動計測に基づいた従来の見解では、ある領野における視覚特徴の神経表象は、その領野内で生成されてその領野における支配的な神経表象になると考えられてきた。
これに対して、まず低次領野において神経表象のプロトタイプ、すなわち「前駆コード」が少数生成され、それが高次領野に送られ、そこで多くのニューロンに広まる、すなわち「増殖」する、という「前駆コード生成→増殖仮説」も立てられている。
この仮説では、大脳皮質の階層的に異なる複数領野にまたがった情報処理によって神経表象が形成される点で、従来の説とは異なった計算原理を提唱する形だ。しかし、このような神経表象の生成過程を解明するためには、個々のニューロンの活動ではなく、複数のニューロンが形成する神経回路を明らかにする必要があり、長年にわたって脳科学の大きな課題となっていた。
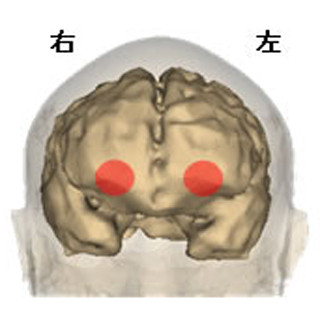
下部側頭葉は、物体の形状に関する情報処理の最終ステップであり、互いに連関した2つの異なる図形(例えば、鉛筆と消しゴム)が1つのまとまった「図形間対連合」として表象されることが知られている。下部側頭葉は、隣接した2つの領野、低次側の「TE野」と高次側の「36野」からなるが(画像2)、TE野の多くのニューロンがそれぞれ個々の図形を表象しているのに対して、36野には図形間対連合を表象するニューロンが数多く存在することが報告されてきた。今回の研究では、図形間対連合の表象が、どちらの領野において、どのような神経回路によって生成されるかを、複数ニューロン活動の同時記録と、それらのニューロン間の機能的結合を解析することによって明らかにした。

|
|
画像2。下部側頭葉における図形間対連合の表象。(上)マカクザルの脳を横から見たもの。右が前側、左が後ろ側。(下)上の点線のレベルで切った脳の断面図。左が外側、右が内側 |
今回の研究では、提示された手がかり図形に対して特定の対図形(画像3)を思い出す「視覚性対連合記憶課題」(画像4)を学習させたマカクザルにおいて、下部側頭葉の隣接した領野であるTE野と36野のそれぞれから課題遂行中に複数ニューロン活動の同時記録を行い、図形間対連合の神経表象を生成する神経回路が調べられた。神経回路を調べるために、「神経活動間の相互相関」(画像5・6)を計算することにより、活動を同時記録したニューロン群について、どのニューロンからどのニューロンへと情報が送られているのかが調べられた。
視覚性対連合記憶課題についてもう少し説明すると、幾つかの対となる事柄(例えば、鉛筆と消しゴム)をあらかじめ憶えてもらい、対の内の片方(例えば、鉛筆)を提示して、もう片方(消しゴム)を思い出してもらうという記憶課題。側頭葉に損傷を持つ患者は、この課題の成績が著しく低いことが知られている。
また神経活動間の相互相関とは、2つのニューロンの活動を同時に記録している時、互いの発火タイミングに相関関係があるかどうかを見ることによって、ニューロン間の機能的結合を推定する方法のこと。例えば、ニューロン1が、ニューロン2よりも数ミリ秒だけ遅れて発火する確率が高い場合、ニューロン2からニューロン1に向けた機能的結合があることが考えられる。つまり、ニューロン2が情報の「送り手」であり、ニューロン1が情報の「受け取り手」である可能性があるというわけだ。反応選択性が特定されたニューロン同士の活動の相互相関を調べることによって、何を表象するニューロンから何を表象するニューロンへと情報が送られているのかがわかる。

|

|
|
画像3(左):サルが視覚性対連合記憶課題のために学習した6組の図形対。画像4:視覚性対連合記憶課題。学習したサルがレバーを引くと、モニターに手がかり図形が提示され、想起期間の後、選択図形が提示される。サルが手がかり図形の対図形(ここでは、左下の選択図形)を正しく選べば正解だ |
|

|

|
|
対連合記憶課題を遂行中のサルのTE野から同時記録されたニューロン対の例。画像5(左):TE野で同時に記録された2つのニューロンの反応を表したもの。ニューロン1(上)は最適刺激とその対図形に選択的に反応し、ニューロン2(下)は最適刺激に対してのみ反応している。画像6:これらのニューロン間の機能的結合を表す相互相関ヒストグラム。横軸は、ニューロン2のスパイク時刻を0とした時のニューロン1の発火タイミング、縦軸はその発火イベントが起こった確率を表している。灰色の横線は、統計的有意性の閾値だ。横軸の0よりも右側にピークがあることから、ニューロン1の発火がニューロン2の発火に遅れて起こる、つまり、ニューロン2が情報の送り手であり、ニューロン1が情報の受け手であることがわかる |
|
その結果、低次側の領野であるTE野には、個々の図形を表象するニューロンから図形間対連合を表象するニューロンへと情報を送る神経回路が数多く存在することがわかった(画像7~9)。これは、対連合表象の「前駆コード生成」過程を表していると考えられる(画像11左上)。
画像11は、下部側頭葉における図形間対連合の表象に至るメカニズムのモデルだ。ニューロン集団による対連合表象の強さは、TE野よりも36野の方が高い(下のグラフ)ことから、対連合表象は36野で生成されると考えられてきた。今回の研究において、それぞれの領野の神経回路を調べることにより、図形間対連合の表象は、まずTE野において個々の図形の表象ニューロンからの収束的な結合(画像11の左上)によって「前駆コード」が生成され、それが36野に運ばれて、図形間対連合の表象ニューロン同士の結合(画像11の右上)を介して「増殖」することによって、支配的な神経表象となることが示されたというわけだ。

|

|
|
TE野の神経回路における図形間対連合表象の「前駆コード」の生成。画像7(左)は、TE野において方向性のある機能的結合を示したすべてのニューロン対の中で、情報の受け手側(上)と、送り手側(下)とそれぞれ判定されたニューロンの反応を表したもの。TE野では、送り手側のニューロンに比べ、受け手側のニューロンの方がより強く図形間対連合を表象していることがわかった。このことから、TE野では個々の図形を表象するニューロンからの収束的な結合(画像8)によって、図形間対連合表象の「前駆コード」が生成されることが示唆された |
|

|

|
|
図形間対連合表象は、TE野で「前駆コード」が生成され、36野で「増殖」する。左右のグラフは、それぞれTE野(画像9(左))と36野(画像10)において、方向性のある機能的結合を示したすべてのニューロン対について、送り手側と受け手側におけるそれぞれのニューロンの図形間対連合の表象の強さを表したもの。TE野では送り手側よりも受け手側の方が図形間対連合をより強く表象しているのに対して、36野では送り手側も受け手側も同程度に強く図形間対連合を表象していることが判明。このことから、図形間対連合の表象はTE野においてその「前駆コード」が生成され、36野においてそれが「増殖」することが示唆された |
|

|
|
画像11。下部側頭葉における図形間対連合の表象に至るメカニズムのモデル |
一方、高次側の領野である36野では、そのような神経回路は見られず(画像10)、代わりに、図形間対連合を表象するニューロン同士が結合し、情報を送る側のニューロンが先に対連合表象を示し、情報を受け取る側のニューロン集団がやや遅れて相互結合を強める過程と並行して対連合表象を示すようになることがわかった(画像12)。
これは、対連合表象の「増殖」過程を表していると示唆される(画像11右上)。以上の結果から、視覚特徴の表象は、従来の見解とは異なり、その表象が支配的である領野よりも低次の領野において少数の「前駆コード」が生成され、それが高次領野における「増殖」過程を経ることによって、支配的な神経表象となることが示された。
画像12は、36野の神経回路における対連合表象の「増殖」過程。36野において、方向性のある機能的結合を示したすべてのニューロン対について、対連合表象の強さの経時変化を表したものだ。横軸の時間は、送り手側のニューロンの対連合表象の強さが上昇し始めた時刻を0としている。送り手側のニューロンの対連合表象の強さが上昇した後、受け手側のニューロンの対連合表象の強さが遅れて上昇してくることから、36野における対連合表象の「増殖」過程を表していると考えられるという。

|
|
画像12。36野の神経回路における対連合表象の「増殖」過程 |
今回の研究は、隣接した2つの領野における神経回路を解析、比較することにより、「外界の内部表現」が大脳皮質内の情報データベースとしてどのように生成されるかについて従来の定説とまったく異なる新しい計算原理「前駆コード生成→増殖仮説」を実証した。これにより、私たちの脳が世界を表象する原理についての理解が深まるのみならず、階層的な構造を持つ人工データベースの効率的設計にも役立つと期待される。また、この内部表象は、外界の認識を媒介するのみならず、内部的に蓄積された長期記憶の表現を担っている記憶データベースであることも知られている。今回の研究で示されたように、低次領野における少数の「前駆コード」を選択的に制御することによって、高次領野の表象に大きな影響を与えられる可能性があることから、将来は内部表象が媒介する認知過程・記憶過程に関わるさまざまな神経疾患の治療法につながる可能性も期待される。