東京大学などは6月16日、受精に関わる精子融合因子「IZUMO1」と卵子受容体「JUNO」の認識機構を解明したと発表した。
同成果は、東京大学大学院薬学系研究科 大戸梅治准教授、石田英子特任研究員、清水敏之教授、福島県立医科大学医学部附属生体情報伝達研究所 井上直和准教授、大阪大学大学院工学研究科の内山進准教授らの研究グループによるもので、6月15日付けの英国科学誌「Nature」に掲載された。
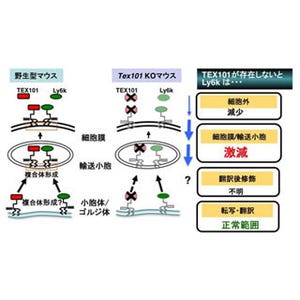
哺乳類の受精では、まず精子が卵子に接着し、その後両者の膜融合が起こることでそれぞれの遺伝情報が組み合わされ、遺伝的に新たな個体が作られる。IZUMO1とJUNOは、それぞれ精子表面と卵子表面に存在するタンパク質で、両者が複合体を形成することで精子は卵子表面に接着する。いずれのタンパク質も受精において必須であることは知られているが、どのような相互作用で結合しているのかは不明となっていた。
今回、同研究グループは、IZUMO1とJUNOの相互作用様式をX線結晶解析によって明らかにするために、それぞれの細胞外領域を大量に調製することに成功。得られたタンパク質を結晶化し、2.0~3.2Åの分解能でIZUMO1単体、JUNO単体、IZUMO1-JUNO複合体の3種類の結晶構造を明らかにした。
この結果、IZUMO1は細長いロッド状の構造、JUNOは球状の構造をしていることがわかった。また、IZUMO1とJUNOは1対1の比率で複合体を形成しており、基本的にそれぞれ単体の場合と同じ構造となっていた。
IZUMO1は、中央のβヘアピン構造が主にJUNOとの結合に関与していて、この部分はN末端側IZUMOドメインとC末端側イムノグロブリン様ドメインとがジスルフィド結合でつながれることにより安定化されていた。
一方JUNOは、葉酸受容体(FRs)と非常によく類似しており、FRsにおける葉酸が結合する部位に相当する疎水性ポケットを持っていた。しかし、JUNOは葉酸と結合しないことが知られている。これは葉酸の認識に関与するいくつかのアミノ酸残基がJUNOでは保存されていないことに加えて、ポケット内部の側鎖がポケットを狭める形で配向しているためであることが、構造から明らかになった。

|
IZUMO1とJUNOの構造。(a)IZUMO1のドメイン構成(上)と構造(下)。(b)JUNO(上)とFRβに葉酸が結合した構造。水色がJUNO、緑色がFRβ、黄色が葉酸 |
またIZUMO1-JUNO複合体は、IZUMO1中央のβヘアピン構造とJUNOの疎水性ポケットの裏側で結合しており、互いに相補的な形状で相互作用していた。特に互いのトリプトファン残基が相互作用界面に存在し、これらのトリプトファン残基は他の生物種でも保存されており、基本的な結合様式は共通していると考えられる。実際に、結合に重要だと考えられるIZUMO1の残基に変異を入れた変異体IZUMO1を発現させた細胞は卵母細胞への接着が著しく減少したという。

|
IZUMO1-JUNO 複合体の構造。(a)IZUMO1-JUNO複合体の全体図。(b)IZUMO1-JUNO複合体の相互作用界面の拡大図。左図はJUNO側の表面図(シアン)、右図はIZUMO1側の表面図(緑、グレー、紫) |
同研究グループは、これらの構造情報をもとに、IZUMO1とJUNOの結合を阻害するような新しい作用機構の避妊薬の開発につながることが期待されると説明している。