科学技術振興機構(JST)、同志社大学、ペプチドリーム、理化学研究所(理研)の4者は10月9日、アルツハイマー病の原因と考えられる「アミロイドβタンパク質(Aβ)」を産生する酵素の特性を解明し、新しいAβ産生抑制方法を開発したと共同で発表した。
成果は、同志社大大学院 生命医科学研究科の舟本聡准教授、同・大学院 脳科学研究科の井原康夫教授、同・大学 生命医科学部医生命システム学科の西川喜代孝教授、同・高橋美帆助教、ペプチドリームの佐々木亨主任研究員、理研 神経蛋白制御研究チームの西道隆臣チームリーダー、同・斉藤貴志副チームリーダーらの共同研究チームによるもの。研究はJST課題達成型基礎研究の一環として行われ、詳細な内容は日本時間10月9日付けで英オンライン科学誌「Nature Communications」速報版に掲載された。
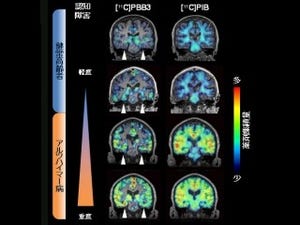
日本の認知症患者数は460万人にのぼると推計され、発症の前段階のいわゆる予備群も400万人ほど存在することが最近になってわかってきた(平成24年厚労省調べ)。その中でもアルツハイマー病は認知症の7割近くを占め、このままでは大きな社会的損失を生み出すものと懸念されている。この傾向はほかの先進諸国にも見られ、アルツハイマー病の克服はもはや世界的重要課題といっても過言ではない状態だ。
アルツハイマー病は、脳内のAβの凝集、蓄積が原因で発症する「アミロイド仮説」が現在のところ主に唱えられている。まずAβの蓄積や細胞毒性が生じて、それが原因となり「タウタンパク質」の凝集と毒性を引き起こし、結果的に神経細胞が機能不全となってアルツハイマー病が発症するというものだ。
Aβは、「βセクレターゼ」と「γセクレターゼ」という2種類のタンパク質分解酵素が、2段階に「アミロイド前駆体タンパク質(APP)」の切断を触媒することで(前者が第1段階の「β切断」を、後者が第2段階の「γ切断」をそれぞれ触媒)産生される40個のアミノ酸からなるタンパク質断片である(画像1)。約40個のアミノ酸から構成されるペプチドだが複数あり、中でも42個のアミノ酸からなる「Aβ42」は、「Aβ40」よりも毒性や凝集性が高く、これが真の原因と現在では考えられている。
従って、アルツハイマー病の予防・治療の最も効果的な方法は、これら2つの酵素活性を抑制することと考えられているが、これらの酵素は生体内で重要なタンパク質分解も担っているため、単なる酵素活性の阻害や酵素の欠損では、生体に重篤な障害を引き起こしてしまうことがわかってきた。

|
|
画像1。Aβの細胞外領域がβセクレターゼによってβ切断され、膜に残ったC99がγセクレターゼによってさらにγ切断されて、Aβが誕生 |
最近、有望な抗Aβ療法として、毒性の強いAβ42産生だけを抑制する「γセクレターゼモジュレータ」が注目されている。γセクレターゼモジュレータはγセクレターゼの切断部位を変調させる薬剤の総称で、一般には、毒性の高いAβ42の産生を抑制する薬剤を示す場合が多い。γセクレターゼによるC99切断を変調させて、Aβ42をさらに短いAβ38に変換すると考えられている。γセクレターゼによる細胞の分化に必須な「Notchの切断」には影響しないことがわかっている。
このようにまさに理想的に見えるγセクレターゼモジュレータだが、治験では期待した効果が得られないケースや、家族性アルツハイマー病変異を持つγセクレターゼにはほとんど効果がないこと、アルツハイマー病脳のγセクレターゼではAβ42の産生抑制能が高くないことなども報告されている。従って、抗Aβ療法に関して新しい発想に基づいた治療・予防戦略が必要だ。そこで研究チームは、γセクレターゼそのものを標的とするのではなく、γセクレターゼが切断する「基質」に着目し、新しい抗Aβ療法の開発に取り組むこととした。なお基質とは酵素によって何らかの影響を受ける物質の総称のことで、今回に限っては両セクレターゼによって直接的に切断されるタンパク質を指す。
研究チームは、Aβ産生だけの抑制を実現させる糸口として、まずγセクレターゼが基質を認識するメカニズムの解明に取り組んだ。さまざまなタンパク質がγセクレターゼの基質となる条件は、(1)「I型膜タンパク質」であること、(2)「細胞外ドメイン」が分解酵素による切断を受けること、が知られている。しかし、さまざまな種類の基質が存在する中で、γセクレターゼがどのように基質を認識するかについてはよく調べられていなかった。
なお、I型膜タンパク質とはその名の通りに膜に存在するタンパク質の1種で、1回膜貫通ドメインを持ち、細胞外側にアミノ末端を、細胞質側にカルボキシル末端を突出されているものの総称。APPもその1つである(画像1)。また細胞外ドメインとは、特に膜タンパク質で細胞外の領域に突出している領域(画像1)のことだ。
APPはセクレターゼによる切断の違いによって、細胞外ドメインが長いC99と短いC83の2種類に分解され、この内C99がさらにγセクレターゼによって切断されるとAβになることがわかっている(画像2)。これらについて、γセクレターゼによる切断効率を調べると、C83の切断効率がC99よりも5倍ほど高く、切断されやすいことが確かめられた(画像3)。

|

|
|
画像2(左):3種類の膜タンパク質についてのN末端における長さが異なるγセクレターゼ基質の1つで、APP由来基質。点線はC末端側の切断部位を示す。画像3(右):各基質の切断効率の比較表。3種類の膜タンパク質において、アミノ末端の短い基質(C83、V1711、R678)の切断効率(Vmax/Km)は、アミノ末端が長い基質(C99、ΔE、M664)のそれよりも約5倍高いことがわかった |
|
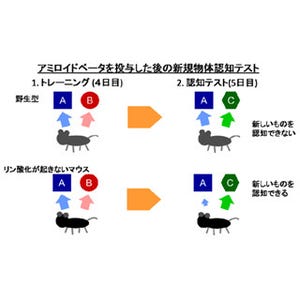
C83はC99よりも細胞外ドメインが短い基質であることから、ほかの代表的な膜タンパク質でγセクレターゼの基質である(Notchと「APP-like protein2(APP様タンパク質2:APLP2)」)についても細胞外ドメインの長さが異なる基質を調製し、切断効率が調べられた(画像4・5)。その結果、これらの分子においても、細胞外ドメインの短い基質で切断効率が約5倍高いことがわかった(画像3)。この結果から、細胞膜内で働くγセクレターゼが基質の細胞外ドメインの長さを識別し、そのドメインの短い方の基質を選択的に切断すると考えられた。なお、APLP2はAPP類似タンパク質の1つ。

|

|
|
画像4(左):。3種類の膜タンパク質についてのN末端のにおける長さが異なるγセクレターゼ基質の1つで、Notch由来基質。画像5(右):同じくAPLP2由来基質。どちらも点線はC末端側の切断部位を示す |
|
さらに、γセクレターゼの基質選択性を検証するために、C99の長い細胞外ドメインをNotchに、Notchの短い細胞外ドメインをC99に交換させたキメラ基質を調製し、γセクレターゼとの結合や切断効率を検討した(画像6)。その結果、Notchの短い細胞外ドメインを持つキメラ型C99はγセクレターゼとの結合が上昇し、さらに切断効率も上昇することがわかった(画像7・8)。一方、C99の長い細胞外ドメインを持つキメラ型のNotchはこの酵素との結合が弱まり、結果的に切断効率も顕著に低下することも判明。これらの結果により、研究チームは、γセクレターゼが基質の細胞外ドメインを認識して、短い細胞外ドメインの基質を効率よく切断することを証明することに成功したというわけである(画像9・10)。

|

|

|
|
画像6(左):細胞外ドメインを交換したC99とNotchのキメラ基質。C99由来領域を黒、Notch由来領域を赤で示す。矢印はC末端側の切断部位を示す。 画像7(中):C99-Notch間キメラ基質のγセクレターゼとの結合と切断。画像3の各キメラ基質とγセクレターゼとの結合が調べられた。 画像8(右):それぞれの基質とγセクレターゼを4℃で保温し、基質に結合しているγセクレターゼ量を検討。細胞外ドメインの短い基質ほどγセクレターゼと顕著に結合している。各キメラ基質の切断。なお、細胞外ドメインの短い基質ほどγセクレターゼにより顕著に切断を受ける |
||

|

|
|
γセクレターゼ基質選択性モデル。 画像9(左):γセクレターゼは基質の細胞外ドメインのN末端側を捕捉すると考えられるという。Notchなどの細胞外ドメインが短い基質のN末端はγセクレターゼからの距離が短いので、この酵素に容易に捕捉され切断される。 画像10:一方、C99などの細胞外ドメインが長い基質のN末端は酵素から離れているので、捕捉されにくく結果として切断を受けにくい |
|
以上の研究によりγセクレターゼが基質の細胞外ドメインを認識して切断することが判明したので、次にC99基質の細胞外ドメインに特異的に結合するペプチド試薬を創製し、C99とγセクレターゼの結合を抑制することにより、Aβ産生だけを抑制する方法の確立を目指した研究が進められた。
「メッセンジャーRNA(mRNA)ディスプレー法」を利用することで、C99細胞外ドメインに特異的に結合するペプチド試薬「C99結合ペプチド」の開発に成功したのである(画像11・12)。実際に試験管内でC99結合ペプチドの効果が検討され、その結果、C99とγセクレターゼの結合を抑制したのに対し、Notchとこの酵素の結合には影響を与えることなく、γセクレターゼによるC99の切断を特異的に抑制することが確認された(画像13・14)。
なおmRNAディスプレー法とは、アミノ酸をコードするランダムな塩基配列を含むDNAライブラリー(1011種類以上)を鋳型に無細胞系翻訳を行い、合成されたタンパク質(またはペプチド)とターゲット間の親和性を利用して、ターゲットに結合するcDNA配列を得る方法。翻訳されるタンパク質はリンカーによりmRNAと結合した状態で維持される。従って、タンパク質は常に自身の遺伝情報を保持しているので、その塩基配列を解析することでスクリーニングで得られたアミノ酸配列を得ることができるという仕組みだ。

|

|
|
C99結合ペプチド。画像11(左):mRNAディスプレーで得られたC99結合ペプチド。画像12(右):共通したアミノ酸配列を赤で示す。AのC99結合ペプチドによる。Aβ産生抑制(B)。ペプチド#1、#2、#4が顕著なAβ産生抑制を示した |
|

|

|
|
C99結合ペプチドによる基質特異的阻害。画像13(左):C99結合ペプチド#4はC99特異的にγセクレターゼとの結合を抑制する。 画像14(右):C99結合ペプチド#4はC99特異的にγセクレターゼによる切断を抑制する |
|
C99はAPPからβセクレターゼによる切断(β切断)で産生されることから(画像1)、C99結合ペプチドのβ切断抑制能についての検討も行われ、C99結合ペプチドはAPP特異的にβ切断をも抑制することがわかった(画像15)。画像15が、C99結合ペプチドによるAPP特異的β切断抑制。C99結合ペプチド存在下で、βセクレターゼとAPPまたはシアル酸転移酵素を反応させて、切断断片産生の影響が調べられた。APPのβ切断はC99結合ペプチドの存在で抑制されたが、シアル酸転移酵素のβ切断には影響を与えなかった。

|
|
画像15。C99結合ペプチドによるAPP特異的β切断抑制 |
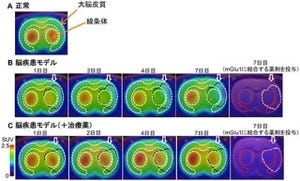
また、C99結合ペプチドを培養細胞に添加しても、Aβ産生を特異的に抑制することがわかり、さらに、C99結合ペプチドを正常なマウスの腹腔に投与した場合も、対照群と比較して有意に脳内Aβ量が低下することもわかった(画像16・17)。以上の結果から、C99の細胞外ドメインを標的としたC99結合ペプチドが、Aβ産生抑制に有効であることがわかった(画像18・19)。

|

|
|
培養細胞・マウスにおけるC99結合ペプチドの効果。 画像16(左):APPとNotchを発現するCHO細胞にC99結合ペプチドを添加して、産生されるAβ量とNotch切断断片(NICD)量が検討された。 画像17(右):C99結合ペプチドは濃度依存的なAβ産生抑制を示したが、NICD産生には影響を与えなかった。Lはγセクレターゼ阻害剤L685458。C99結合ペプチド#4をマウス腹腔に投与(150mg/kg/d)すると、脳内のAβ量が減少した |
|

|

|
|
画像18・画像19:C99結合ペプチドの作用機序。C99結合ペプチドはC99に特異的に結合し、γセクレターゼとの結合を抑制し、その結果として、C99切断(Aβ産生)を抑制する。一方、C99結合ペプチドはNotchなどのほかの分子には結合しないので、これらの基質の切断には影響を与えない |
|
今回の研究により、酵素ではなく基質をターゲットとすることで、基質特異性を維持しながら効果的にAβ産生を抑制することに成功した形だ。しかも、1種類のペプチド試薬で2種類のAβ産生に関わるAPP切断を抑制することができ、まさに抗Aβ療法に一石二鳥の効果をもたらすといえよう。
今回得られたC99結合ペプチドに改良を加えて脳移行性やC99との親和性を高めて、副作用の少ないアルツハイマー病の予防・治療に役立てることが期待されるという。また、このC99結合ペプチドの機能を模倣する低分子化合物のスクリーニングなども視野に入れて、副作用の少ない治療・予防薬の開発に貢献することを目指すとした。
今回の研究で示した阻害剤創製アプローチは、従来のような酵素を標的とする手法ではなく、基質をターゲットとするもので、がんなどのほかの疾患にも適応可能で、副作用の少ない治療・予防薬の開発にも期待ができるとしている。