大阪大学(阪大)は5月31日、ベルギー・ゲント大学との共同研究により、植物の成長に重要な働きをする植物ホルモン「オーキシン」の流れを作るために、植物は細胞内の膜系に存在する2つの因子(「BEN1」と「BEN2」)を利用していることを明らかにし、そのメカニズムは根の形成パターンや葉脈のつながり方などのさまざまな発生過程を制御するのに使われていることも解明したと発表した。
成果は、阪大大学院 理学研究科の田中博和助教、同・北倉左恵子研究員、ゲント大のJiri Friml博士らの国際共同研究チームによるもの。研究の詳細な内容は、米国西部時間5月30日付けで米生物学専門誌「PLOS GENETICS」オンライン版に掲載された。
オーキシンは植物体の中で方向性を持って流れているユニークな成長制御物質で、光や重力の方向に向かって植物が生長する「屈性反応」や、葉や根などの器官の形成などに関わっている植物ホルモンの1種だ。これまでの研究から、オーキシンの流れを作るには「PINタンパク質」が細胞膜に偏って局在することが重要だと考えられている(画像1)。
|
|
|
画像1。PINタンパク質の働きによるオーキシンの輸送の模式図。PINタンパク質が細胞膜で偏って分布し、オーキシンを排出することで組織の中でオーキシンが一方向的に輸送される(緑矢印) |
また、植物の葉や根などの器官が作られる時には、PINタンパク質の偏る方向がダイナミックに変化することも最近の研究でわかってきた。この過程には、PINタンパク質を細胞膜に運んだり、細胞膜から再び細胞内に取り込んで運んだりする、「小胞輸送」のメカニズムが関与していると想像されているが、このPINタンパク質の細胞内輸送に関しては不明な点が多く、詳細はよくわかっていないのが現状だ。
なお小胞輸送とは、タンパク質や脂質などの物質が膜小胞を介して「オルガネラ」やそのほかの細胞内区画、細胞膜の間で輸送されることを指す。細胞膜から小胞が形成され、細胞内に取り込まれることを「エンドサイトーシス」と呼び、逆に細胞内から細胞膜へと小胞が輸送され融合する過程を「エキソサイトーシス」という。
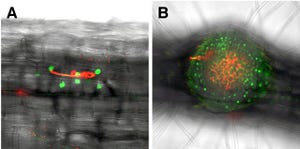
田中助教らはこれまでの研究により、特殊なスクリーニング法を用いて、PINタンパク質の細胞内での動きが異常になったシロイヌナズナの変異体「ben1」と「ben2」を単離していた。今回の研究ではこれら2つの変異体の解析を進め、「ben1;ben2二重変異体」では「初期エンドソーム」を介したPINタンパク質の動きが通常とは異なることが発見された(画像2・3)。
ちなみにben1変異体とは、「ARF GEF類似タンパク質」をコードする遺伝子に変異が入った変異体である。「AEF GEFは低分子Gタンパク質」である「ARF」に作用し、「GDP結合型」のARFを「GTP結合型」に変換する「グアニンヌクレオチド交換因子」だ。ARFによる細胞内輸送経路(特に小胞の出芽)を活性化する因子であると考えられている。
そして初期エンドソームとは、高等植物の細胞中にある、物質代謝やタンパク質の選択的な輸送などの固有の役割を持った。膜で囲まれたオルガネラやさまざまなコンパートメント(区画)の内、細胞の外にある物質や細胞膜の成分が細胞内に取り込まれた時に、初期に到達するコンパートメントのことをいう。また、初期エンドソームは物質を選別し、次の行き先に送る働きがあると考えられている。最近の研究により、シロイヌナズナではトランスゴルジ網(TGN)と同一と考えられるようになったため、「TGN/EE」とも呼ばれる。

|
|
|
画像2(左):初期エンドソーム(TGN/EE)を介した輸送経路の阻害剤ES1を用いた実験。PIN2タンパク質を免疫染色により検出している。ben1変異体とben2変異体はES1に対する応答が野生型(WT)とは異なる。画像3(右):細胞内のタンパク質輸送経路のモデル。BEN1は初期エンドソームからの輸送、BEN2は初期エンドソームへの輸送に関わると考えられる |
|
また、この二重変異体ではPINタンパク質の局在の偏りが減少し、それに伴い胚発生、根の形成、葉脈形成などのパターンが変化することもわかった(画像4~6)。根の形成過程やオーキシンを加えた場合のPINタンパク質の偏りの変化が詳細に調べられ、その結果、二重変異体ではPINタンパク質の局在変化が起きにくくなっていることが判明したのである。
このことは、BEN1、BEN2という因子が、根の形成時の位置情報やオーキシンなどのシグナルに応答したPINタンパク質の再配置に必要であることを示しているという。

|

|

|
|
BEN1とBEN2はPINタンパク質の局在と植物の発生に重要である。 |
||
BEN2タンパク質は膜融合の制御に関わる「Sec1/Munc18ファミリータンパク質(AtVPS45)」で、BEN1タンパク質と同じく、初期エンドソームに存在することが明らかになった。BEN1とBEN2が初期エンドソーム(TGN/EE)を介した輸送に関わるということを併せて考えると、PINタンパク質の再配置にはPINタンパク質を細胞内に取り込み、細胞内輸送経路で選別して次の行き先に運ぶという仕組みが働いていると考えられるという。
BEN2タンパク質が属するSec1/Munc18ファミリータンパク質は膜の融合に関わる因子で、「AtVPS45/BEN2」はこれまでは「トランスゴルジ網(TGN)」という細胞内小器官から液胞へのタンパク質輸送に関わると考えられていた。今回の研究結果から、BEN2は初期エンドソームへの輸送と植物の細胞極性において、極めて重要な働きをしていることが解明された形だ。動物のVPS45類似タンパク質もエンドサイトーシスと細胞極性に関わるものが知られていることから、VPS45が関わる小胞輸送の制御は多細胞生物の組織化に普遍的に関わっている可能性があるとする。
BEN1とBEN2が関わるタンパク質輸送システムは、植物の胚発生、器官形成、葉脈形成などのさまざまな発生過程に利用されており、これらの分子を調節することにより、オーキシンの輸送の方向性を人為的に制御して、植物の形を変化させることが可能になるかも知れないという。今後、これらの分子の作用機構を探求することで、タンパク質の選択的輸送と細胞極性の研究の加速につながると期待されるとしている。