理化学研究所(理研)は12月18日、脳や脊髄の病巣に蓄積するタンパク質「TDP-43」の安定化が、全身の筋肉マヒを起こす神経変性疾患「筋萎縮性側索硬化症(Amyotrophic Lateral Sclerosis:ALS)」の発症時期を決定する要因であることを明らかにしたと発表した。
成果は、理研 脳科学総合研究センター 運動ニューロン変性研究グループの山中宏二チームリーダー、同・渡辺祥司研究員(現・同志社大学高等研究教育機構 助教)らの研究グループによるもの。
研究の詳細な内容は、日本時間12月13日付けで米国の生化学・分子生物学会誌「The Journal of Biological Chemistry」オンライン版で公開され、印刷版2013年2月号に掲載される予定だ。
ALSは、全身の筋肉を支配する運動神経細胞を侵し、呼吸筋を含む全身の進行性の筋肉マヒを引き起こす原因不明の神経変性疾患だ。ALSの約90%は非遺伝性、残りの約10%が遺伝性で、わが国では約8500人のALS患者がいると推定されている(出典:公益財団法人難病情報センター)が、有効な治療法は見つかっていない。患者の苦痛に加え、長期にわたって負担の重い介護を必要とするため、原因の解明と治療法の開発が求められている状況だ。
主に細胞内の核に分布するTDP-43は、細胞質と核との間を往来して、DNAからmRNA(メッセンジャーRNA)を経てタンパク質を合成する転写・翻訳制御やスプライシング制御など多面的なRNA制御を行うと考えられている。
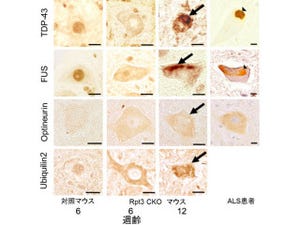
近年になって、ほぼすべてのALSの病巣からTDP-43が核から細胞質へ脱出して異常に凝集・不溶化し、蓄積することが発見された。さらに、非遺伝性や遺伝性ALSの一部からTDP-43遺伝子の変異が30種類以上見つかっていることから、遺伝的な背景の有無に関わりなく、TDP-43の機能異常がALSの病態に関与していることが示唆される(画像1)。
したがって、アルツハイマー病における「アミロイドβ」やパーキンソン病における「αシヌクレイン」といったタンパク質の病的蓄積と同様に、ALSの病巣におけるTDP-43の異常蓄積は重要な要因と考えられるところだ。しかし、TDP-43遺伝子の異常がどのようにALS発症に関わっているのか、その詳細については不明だった。
画像1は、TDP-43の構造とALS患者における変異。上の図のTDP-43は、414のアミノ酸からなり、多くの組織・細胞で恒常的に発現している。2つの核内移行シグナル(NLS #1、#2)および核外移行シグナル(NES #1、#2)の両方を持ち、核と細胞質との間を往来しているという特性を持つ。
また、2つのRNA結合領域(RRM1、2)と、「グリシン(Gly)」が多数並ぶ1つの「グリシンリッチドメイン(Gly rich region)」を有しており、翻訳制御やスプライシング制御などのRNA制御を行うと考えられている。
一方、下の図のALS患者に見られるTDP-43変異は、タンパク質のC末端のグリシンリッチドメイン(274~414アミノ酸)に集中している。特に、G298S、A315T、M337V、Q343R、G348C、N352S、A382Tの7種の変異TDP-43を持つ遺伝性ALS患者数が比較的多い(赤字)。

|
|
画像1。TDP-43の構造とALS患者における変異 |
研究グループは、変異TDP-43の生化学的特徴と、TDP-43遺伝子の変異を持つ遺伝性ALS患者の発症年齢、罹病期間(疾患の進行)のいずれが相関するのかを明らかにするため、19種類のTDP-43遺伝子変異を持つ合計81人の遺伝性ALS患者について臨床情報を収集した。
一方で、マウス神経の培養細胞に正常(野生型)と変異TDP-43を発現させ、タンパク質の細胞内分布、凝集能、タンパク質の半減期を比較・検討し、これらの生化学的特徴とALSの発症年齢または罹病期間との関連を調べたのである。すると、以下のことが明らかになった。
変異TDP-43は、野生型より細胞質に移行しやすく、高い凝集能を持つ傾向があることが判明したが、これらと発症年齢や罹病期間との相関はなかった。また、比較的患者数の多い7種類の変異TDP-43の半減期を、「パルス-チェイス法」により測定を実施(画像2)。
その結果、すべての変異TDP-43の半減期が顕著に長くなったため、変異TDP-43遺伝子がタンパク質の安定化を引き起こすとわかったのである。さらに重要なことは、タンパク質半減期が長くなってタンパク質がより安定化する変異TDP-43遺伝子を持つ患者ほど、早期にALSを発症することが判明した。
画像2は、TDP-43の半減期はALSの発症時期と相関することを示したもの。野生型(WT)、比較的患者数の多い7種の変異TDP-43(G298S、A315T、M337V、Q343R、G348C、N352S、A382T)、核外・核内移行シグナルを破壊した変異体(NES1/2、NLS1/2)それぞれを、マウス神経細胞内で一過的に発現させ、パルス-チェイス法で半減期を決定した(A、B)。その結果、すべての変異TDP-43の半減期が、顕著に長くなることが判明したのである。
さらに変異TDP-43の半減期と、発症年齢(C)または罹病期間(D)との相関も調べられた。その結果、半減期と発症年齢に負の相関(右肩下がりのグラフ)が認められ(C)、半減期の長い変異を有するALS患者ほど平均発症年齢は早くなることがわかったのである。一方、変異TDP-43のタンパク質半減期と罹病期間の間には、相関は確認されていない(D)。

|
|
画像2。TDP-43の半減期はALSの発症時期と相関することを示したもの |
次に研究グループは、TDP-43の安定化がALSの発症時期に関わることに着目し、低分子化合物「Shield1」でタンパク質の安定性を制御できるタグ(DDタグ)を利用して、TDP-43の異常蓄積を再現するマウスの神経細胞モデル「DD-TDP-43」を作製(画像3-A)。
Shield1の添加により、TDP-43を安定化させると、時間に依存してTDP-43の切断が見られ、界面活性剤(Sarkosyl)に溶けない凝集体の増加も見られた(画像3-B)。これらの現象は、ALS患者の病巣で見られるTDP-43の生化学的特徴を再現している。そしてTDP-43の安定化・蓄積により、顕著に神経細胞の細胞死が誘導された(画像3-C)。
画像3は、TDP-43を安定化する細胞モデル。(A)ではTDP-43の末端に、FK506結合タンパク質の変異体(DD)を結合させた融合タンパク質に、Shield1を加えるとTDP-43が安定化する(+Shield1)。一方、Shield1を加えないでいると不安定化し、急速に分解される(-Shield1)結果を示したもの。
(B)はTDP-43をShield1により任意の時間安定化させた後、細胞を3種の異なる界面活性剤を用いて段階的に溶解させ、Sarkosylに対する不溶性を検討したもの。この結果、安定化の時間に依存してSDSにより溶化されるTDP-43量(赤枠)が増加した。つまり、Sarkosylでは、不溶化のままのTDP-43や切断されたタンパク質断片が増加したことを示す。以上の結果は、ALS患者の病巣で見られるTDP-43の生化学的特徴を再現している。
そして(C)から、安定化したTDP-43が細胞内で蓄積すると死細胞数(PI陽性細胞)の増加が見られ、安定化したTDP-43は細胞毒性を発揮することがわかった。

|
|
画像3。TDP-43を安定化する細胞モデル |
また安定化したTDP-43は、切断されたり不溶化したりするだけではなく、TDP-43自身のmRNAを分解してタンパク質の異常な蓄積を防ぐ機能(mRNAの負の制御能)が低下することも判明。さらに、細胞内の主要なタンパク質分解装置である「プロテアソーム」の活性を、安定化したTDP-43が顕著に阻害することもわかった(画像4)。
画像4は、安定化させたTDP-43の蓄積による、細胞内のプロテアソーム活性の低下示す証拠。細胞内のプロテアソーム活性を検討するために、緑色蛍光タンパク質の1種の「AcGFP」の末端に分解する目印となる「ユビキチン変異体(UbiquitinG76V)」を結合させた。この融合タンパク質は細胞内でプロテアソームにより分解されるため、プロテアソームの活性は低下しているとAcGFPタンパク質が蓄積し、その量が増加する(A)。このユビキチン-AcGFP複合体とDDタグを結合したTDP-43をマウス神経細胞に一過的に発現させ、Shield1による任意の時間のTDP-43の安定化が施された。
次に、タンパク質を分離してサイズや量を解析する「ウエスタンブロット法」により、抗AcGFP抗体でAcGFP量を検出・定量を実施(B、C)したところ、AcGFP融合ユビキチン変異体の量は、安定化したTDP-43の蓄積により増加することがわかった(B上段、C)。この増加はプロテアソーム阻害剤(MG132)を添加した時と同程度であったことから(C右)、TDP-43の蓄積はプロテアソーム活性の低下を引き起こすことが判明したのである。

|
|
画像4。安定化させたTDP-43の蓄積による、細胞内のプロテアソーム活性の低下を示す証拠 |
以上から、遺伝性ALS患者の臨床情報とTDP-43の生化学的特性の相関を検討し、TDP-43の安定化がALSの発症時期を決定する要因であることが確認された次第だ。
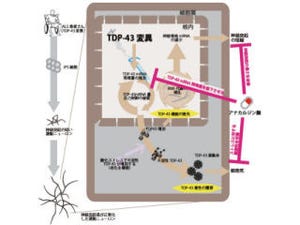
そして安定化したTDP-43は、半減期の延長、自己mRNAの負の制御能の低下、およびプロテアソーム活性の低下を引き起こすことも判明。その結果、慢性的に細胞内で蓄積しやすい環境が誘導され、TDP-43が蓄積すると運動神経細胞に対して毒性を発揮し、運動神経細胞に細胞死を誘導することがわかったのである(画像5)。
画像5は、今回の研究内容をまとめための。変異の導入または未知の因子によりTDP-43が安定化すると、TDP-43の切断・不溶化が起こる。そこへ、TDP-43の持つ自己mRNA制御能の喪失や、プロテアソーム活性の低下が続く。これにより、慢性的にTDP-43の発現量が上昇し、より一層、細胞内でTDP-43が蓄積しやすい環境となり、最終的に運動神経細胞の細胞死を誘導すると考えられるというわけだ。

|
|
画像5。今回の研究内容のまとめ |
研究グループによれば、今回の成果は、遺伝性ALSだけでなく非遺伝性ALSの発症にも極めて重要な要因であることが示唆されるという。今後、開発された細胞モデルを利用して、TDP-43の安定化により運動神経細胞死に至る詳しい機序を理解し、ALS発症メカニズムの解明や治療薬の開発が進展することが期待できるとしている。